mRNA expression levels of cGAS in pan-cancer
To explore the role of cGAS in cancer, the transcription levels of cGAS were investigated based on TCGA datasets. Relative to normal tissues, the mRNA expression levels of cGAS were significantly overexpressed in the majority of the malignant tumor types (Fig. 1), with the exception that there was no significant difference in PAAD, SKCM and UCEC. Contrarily, it was significantly underexpressed in PRAD. For LGG and OV, difference analysis was not performed as no normal samples were not provided in TCGA. We further assessed cGAS expression according to the American Joint Committee on Cancer (AJCC) cancer stages, the results revealed that there was no significant difference in cGAS levels between different stages in most cancers (Supplementary Fig. 1).
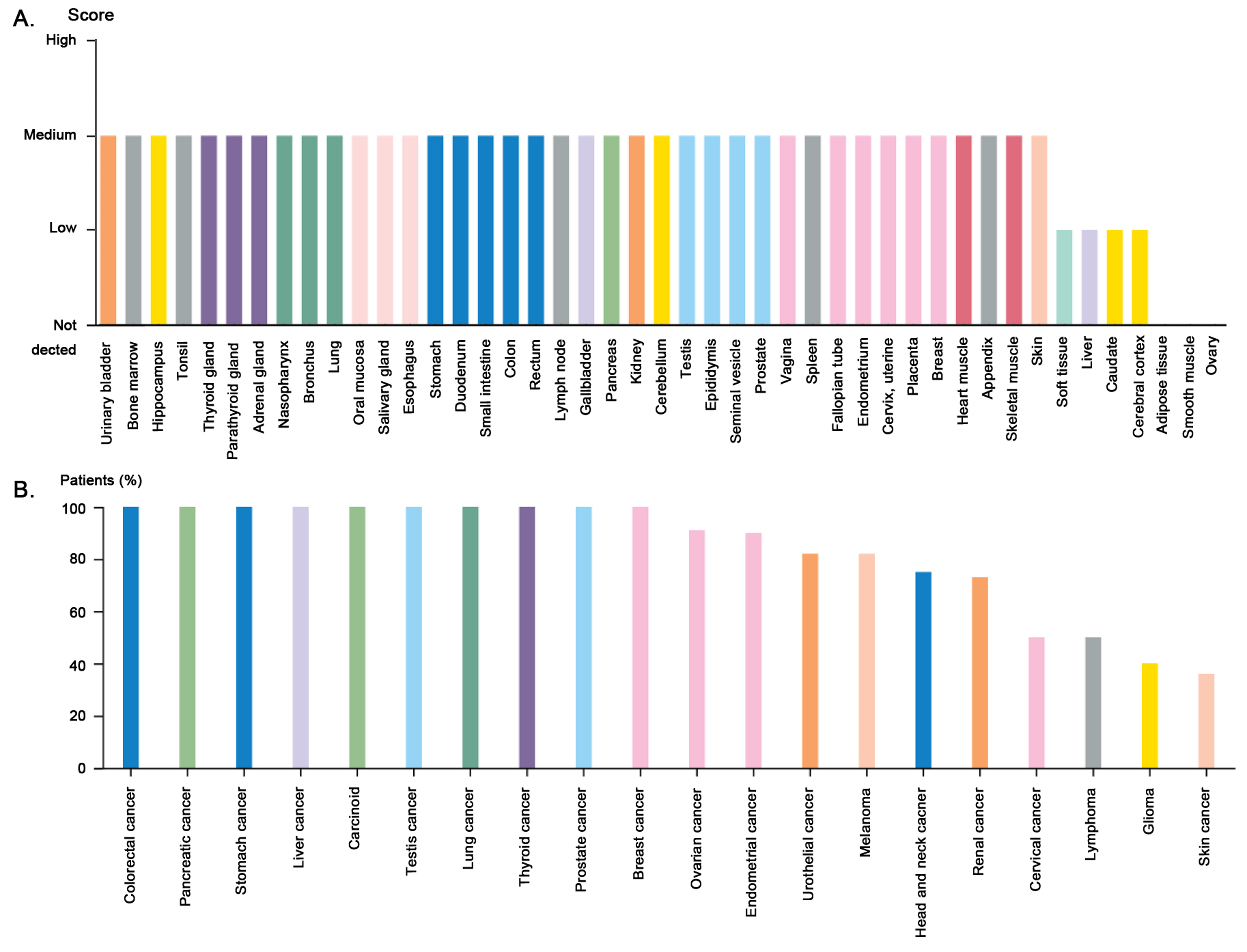
The protein levels of cGAS were analyzed based on the Human Protein Atlas (HPA) database (Supplementary Fig. 2). We found that moderate cytoplasmic and membranous staining of cGAS was observed in most normal tissues. Similarly, a majority of cancer tissues displayed moderate to strong cytoplasmic staining with additional membranous positivity in several tissue.
Genomic alterations of cGAS in pan-cancer
To understand the potential mechanisms of the abnormally expression of cGAS in pan-cancer, we also analyzed the copy number variation (CNV), mutation and methylation data of cGAS from TCGA and CCLE. The CNV and mutation data of TCGA were analyzed using cBioPortal. As shown in Fig. 2A, CNV of cGAS were observed in most of cancers, among which PRAD displayed the highest deep deletion frequency. While CNV of cGAS were not found in GBM, LGG, KIRP and THYM. Across the 20 cancer types, samples harboring cGAS deletion exhibited lower mRNA expression than those harboring diploid cGAS, while mRNA levels were higher in these harboring cGAS overexpression or gain than these exhibit diploid cGAS (Fig. 2C, Supplementary Fig. 3). Consistently, correlation analysis revealed that copy number values were positively correlated with cGAS mRNA expression in most of cancers (Fig. 2D, Supplementary Fig. 4). Unlike CNV, the frequency of cGAS mutation was quite low ranging from 0 to 2.5% (Fig. 2B). Across the 20 cancer types, UCEC exhibited the highest mutation frequency, whereas no mutations were found in GBM, LGG, KIRP and THYM. Besides, samples harboring mutated cGAS exhibited higher mRNA expression than those harboring wild-type cGAS (Fig. 2E).
We further analyzed the methylation levels of cGAS promoter by analyzing the MethHC database. The results revealed that the methylation levels of the cGAS gene promoter were significantly downregulated in CESC, COAD, KIRC and KIRP, but that it was significantly upregulated in LIHC, LUSC, PRAD and READ (Supplementary table 3, Supplementary Fig. 5). We also performed correlation analysis between methylation and mRNA expression of cGAS using cBioPortal. As shown in Supplementary Fig. 6, there was a reverse relationship between mRNA expression and methylation levels in all of the 20 cancer types.
To further understand the genomic alterations of cGAS, we also analyzed the data of cancer cell lines from CCLE using cBioPortal. The CNV and mutation frequency of cGAS in cancer cell lines were high relative to that in human cancers, both ranging from 0 to about 20% (Supplementary Fig. 7A and 7B). In line with the results of TCGA, copy number values were positively correlated with cGAS mRNA expression (Supplementary Fig. 7C). In addition, we found that there was a positive relationship between mRNA expression and protein levels (Supplementary Fig. 7D). Whereas, no significant difference was found in copy number values between cells harboring mutated cGAS and these with wild-type cGAS (Supplementary Fig. 7E).
Association between cGAS expression and interferons signaling in pan-cancer
We examined the correlations between cGAS expression and interferons. As shown in Fig. 3A, cGAS expression positively correlated with most type I IFNs but not type II IFNs in most cancers. Whereas there were no significant correlations of cGAS to type I or type II IFNs in GBM.
Correlation of cGAS expression with signatures of immune cell infiltration
To investigate whether cGAS was involved in the regulation of immune response to these cancers, we explored the correlations between cGAS expression and immune cell infiltration using the CIBERSORT database. As shown in Fig. 3B and 3C, cGAS levels were positively correlated with the degree of CD8 T cells infiltration in 6 cancer types, including BRCA, HNSC, KIRC, LGG, LUAD and STAD. Contrarily, cGAS levels was negatively correlated with CD8 T cells in UCEC. Of memory CD4 T cells, cGAS levels were positively correlated with activated memory CD4 cells in 12 cancer types. Correlations between cGAS expression and resting memory CD4 T cells were significantly in 6 cancer types, including 3 negative correlations (LUAD, OV and STAD) and 3 positive correlations (LGG, PRAD and UCEC). Besides, cGAS levels were positively correlated with TFH in 12 cancer types, and negatively correlated with Treg in 5 cancer types. Among cell types related to innated immunity, cGAS levels were positively correlated with the degree of infiltration of Macrophages M1 and activated DCs in14 and 8 cancer types, respectively. Of Macrophages M2, there were negative correlation in 8 cancer types and positive correlation in 1 cancer type (CRC). cGAS levels were negatively correlated with resting and activated mast cells in 11 and 3 cancer types, respectively. Contrarily, cGAS levels was positively correlated with activated mast cells in GBM. Besides, cGAS levels were positively correlated with neutrophils in 8 cancer types. cGAS levels were significantly correlated with activated NK cells in 6 cancer types, including 5 positive correlations (BLCA, CRC, HNSC, KIRC and STAD) and 1 negative correlation (LIHC).
To further investigate the correlation between cGAS expression and immune cells infiltration, we also examined the associations between cGAS levels and the expression of immune marker genes of various immune cells[25, 26]. Firstly, we analyzed the different functional T cells, including T cells, CD8 T cells, cytotoxic cells, Th1 cells, exhausted T cells and Treg (Fig. 4A). cGAS expression significantly correlated with most immune marker genes in 11 cancer types, except for GBM. In GBM, cGAS levels were significantly correlated with only 13 genes. Next, cells related to innate immunity were analyzed, including DCs, tumor-associated macrophages (TAMs), M1, M2, mast cells, neutrophils and NK cells (Fig. 4B). The result revealed that cGAS expression significantly correlated with most immune marker genes in cancers analyzed, except for CESC, ESCA, GBM, OV and STAD.
Relationship between cGAS levels and DDR genes
Studies reported that cGAS involved in DNA damage repair, thus, we investigated correlations between cGAS expression and DNA damage response and repair pathway gene sets, focused on nonhomologous end joining (NHEJ) and homologous recombination (HR). As shown in Fig. 5A and 5B, cGAS expression significantly correlated with most NHEJ and HR genes in most cancers, except for CESC, ESCA, GBM, and LUSC.
Association between cGAS expression and TMB, MSI and tumor immune microenvironment in pan-cancer
We next examined the associations between cGAS expression and TMB (Fig. 5C). cGAS expression were positively correlated with TMB in BLCA, BRCA, CRC, LGG, LUAD, OV and STAD. Contrarily, cGAS levels were negatively correlated with TMB in LIHC and PRAD. We also examined the correlations between cGAS levels and MSI (Fig. 5D). The results showed that cGAS expression were positively correlated with MSI in BRCA, CRC and STAD, yet, cGAS levels were negatively correlated with MSI in PAAD and SKCM. Besides, the fraction of stromal and immune cells was assessed using ESTIMATE algorithm. The analysis revealed that cGAS expression were significantly correlated with at least one of the immune scores and stromal scores in all of the cancer analyzed (Supplementary Figs. 8 and 9).
Prognostic value of cGAS in pan-cancer
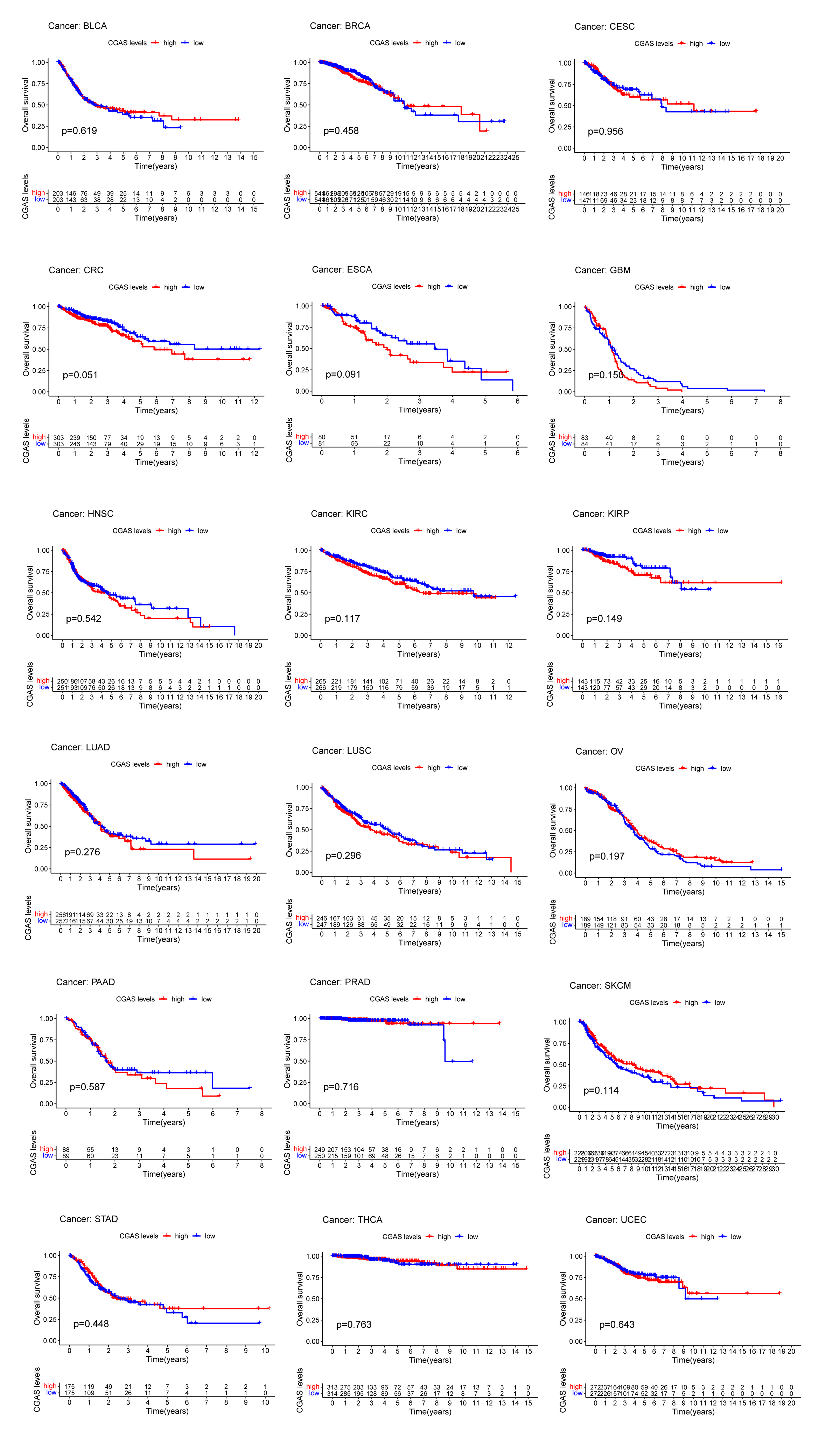
We also determined the prognostic value of cGAS in cancer patients using different databases. In TCGA database, Kaplan-Meier survival curves showed overexpression of cGAS is associated with poor prognosis in LGG (Fig. 6A) and LIHC (Fig. 6B). Whereas, the expression of cGAS failed to predict prognosis in other cancer types (Supplementary Fig. 10). COX analysis showed cGAS was a detrimental prognostic factor in KIRC, KIRP, LGG, LIHC and PAAD (HR > 1, P < 0.05, Fig. 6C). Contrarily, cGAS played a protective prognostic factor in SKCM and STAD (HR < 1, P < 0.05, Fig. 6C). To further examine the prognostic potential of cGAS in different cancers, Kaplan-Meier plotter database was used to assess the association between cGAS expression and patients OS. cGAS had a detrimental effect on esophageal adenocarcinoma, KIRC, KIRP, LIHC, LUSC and PAAD (P < 0.05, Fig. 7A-7F). In contrast, cGAS had a protective effect on OV, PCPG and STAD (P < 0.05, Fig. 7G-7I). Finally, the impact of cGAS expression on patient survival was evaluated using the PrognoScan database. Detailed results are summarized in Supplementary Table 4. cGAS played a detrimental prognostic effect in the lung adenocarcinoma and transitional cell carcinoma (HR > 1, P < 0.05).
Knockdown of cGAS expression inhibits the DNA damage repair signaling pathway and interferon signaling
Finally, we also validated the role of cGAS in DNA damage repair signaling and interferon signaling in NSCLC (Fig. 8, Supplementary Fig. 11). We used small interfering RNA (siRNA) to silence the expression of cGAS. The efficiency of siRNA was validated in RT-PCR assay (Fig. 8A). After knocking down the expression of cGAS, the expression of p-BRCA1, p-CHK1, p-CHK2 and Rad51 was significantly lower level than that in the NC-siRNA cell line (Fig. 8B and 8C). RT-PCR further confirmed the results of western (Fig. 8D). Besides, the expression patterns of γH2AX were significantly higher level than that in the NC-siRNA cell line (Fig. 8C). Furthermore, we also detected the protein level and mRNA level of IFNβ by ELISA and RT-PCR, respectively. The results showed knockdown of cGAS significantly decreased the level of IFNβ (Fig. 8D). Collectively, those results demonstrated that cGAS positively regulate the DNA damage repair signaling and interferon signaling in NSCLC.
{kind=link}
{kind=link}