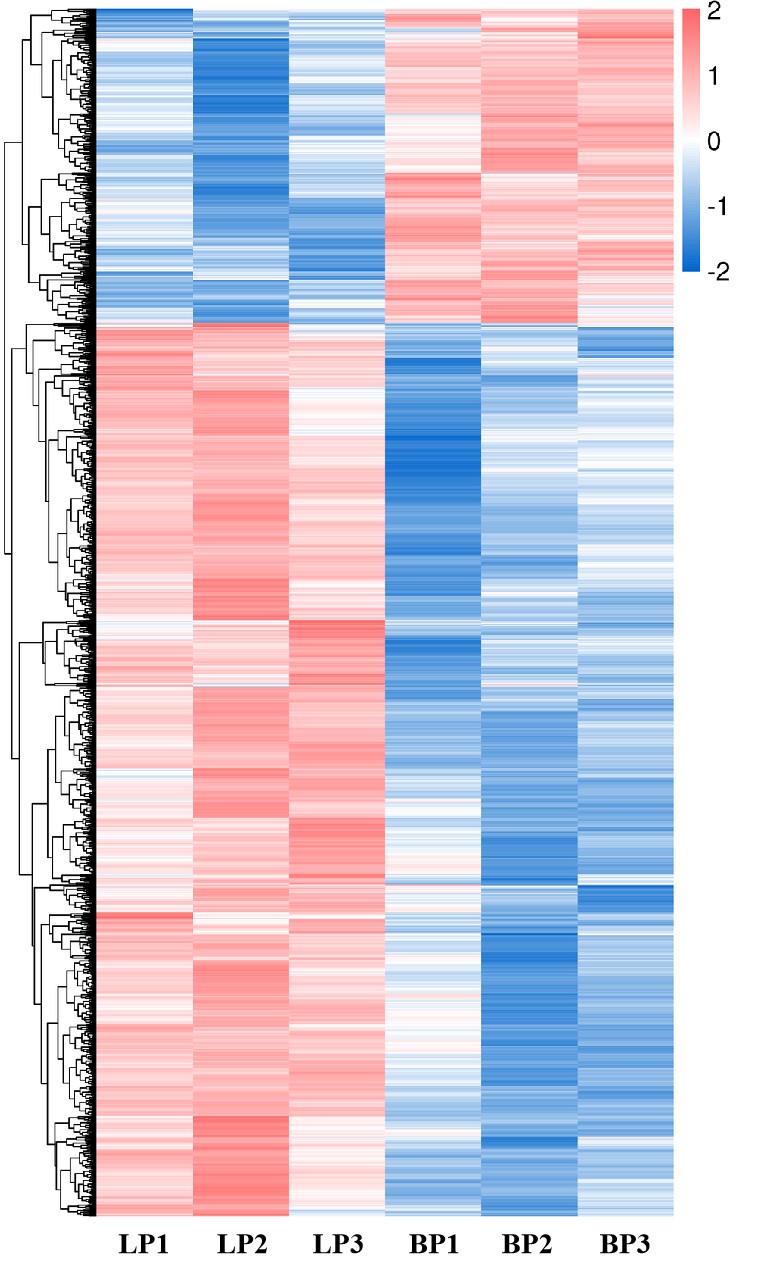
Broodiness is known to be a maternal behavior of poultry, and it is closely associated with HPGA. During the transition from egg-laying to incubation, the transcriptome changes of avian ovaries, however, need to be further explored. In this study, we analyzed the ovarian transcriptome of Changshun hens at LP and BP, and further identified the candidate genes and signal pathways related to broodiness through KEGG and GO enrichment analysis. This study not only provides novel insights into understanding of avian broodiness, but also contributes valuable information to improve Changshun chicken breeding with low nestability in the future.
Considering the fact that avian ovary contains follicles at distinct developmental stages including cortical follicles, white follicles, yellow follicles, and preovulatory follicles, it is therefore deemed to be an ideal model to study the mechanisms of follicular development [30, 31]. Broodiness in avians is generally accompanied by the atresia of follicles and the degradation of reproductive performance. In the present study, we identified several critical signaling pathways related with follicle development in chicken ovaries, including focal adhesion, MAPK signaling pathway, FOXO signaling pathway, and Wnt signaling. The transcriptome profile of geese revealed that ovarian development-related focal adhesion and ECM-receptor interaction were the top two pathways enriched with the most DEGs in incubating ovaries, indicating the potential central roles of these two pathways in the entire ovulation cycle [32]. Similarly, focal adhesion is found to play important roles in prehierarchal follicles of laying and incubating geese [33]. In chickens, focal adhesion is observed to be closely associated with egg production and the process of follicle selection [34, 35]. These findings indicates an important role of focal adhesion in regulating avian ovarian function and egg production. MAPK signaling pathway is one of the most important pathways in ovaries associated with avian age at first egg and egg production [36–38]. MAPK signaling pathway is demonstrated to be involved in cell proliferation and progesterone secretion of granulosa cells from the prehierarchical follicles in chickens [39–41]. Additionally, it is suggested that the granulosa layer within chicken follicles remains undifferentiated and steroidogenically inactive due to the inhibitory actions of MAPK and/or protein kinase C signaling [42]. Wnt pathway is known to be an evolutionarily conserved signaling pathway. In humans, it has been shown to play a pivotal role during human follicle formation and follicle maintenance [43]. In domastic waterfowl, Wnt signaling pathway might be implicated in the follicular development [17, 33, 44]. It was considered to be one of the most important signaling pathways in regulating broodiness of Muscovy ducks [17]. In accordance to these findings, our results further indicated the important role of Wnt signaling pathway in chicken ovaries during broodiness. It was previously suggested that FOXO signaling pathway in ovaries had important functions in the regulation of broodiness in ducks [15]. In the present study, we identified 20 DEGs, including PRKAB2, IGF1 and PTEN, classified into FOXO signaling pathway. PRKAB2 gene is deemed to be associated with live-weight, carcass-weight, leg-muscle-weight and abdomen-fat-weight [45, 46]. IGF1 and PTEN has been reported to be potential key genes that regulate ovulation of ducks [47]. In chickens, IGF1 as an autocrine/paracrine regulator of follicular growth and differentiation is involved in the regulation of follicular development [48–50]. Meanwhile, it stimulates the release of progesterone and affects egg production of chickens [51]. PTEN is the phosphatase of phosphatidylinositol (3,4,5)-trisphosphate (PIP3), which regulates cell proliferation cycles and inhibits cell migration. PTEN is observed to involve in the ovarian function remodeling of laying hens [52]. We found that both IGF1 and PTEN were down-regulated in incubating ovaries in the present study, indicating that they might have critical roles to play in the regulation of ovarian function in incubating chickens.
Vascular smooth muscle contraction, ECM-receptor interaction and GnRH signaling pathways are classical signaling pathways associated with follicle development. Notably, vascular smooth muscle contraction, ECM-receptor interaction, and GnRH signaling pathways in the present study were all down-regulated in the ovaries of incubating chickens. Development of ovarian follicles generally requires the coordinated interactions between theca cells, oocytes and granulosa cells. Transcriptome analysis of circRNA and mRNA in theca cells from different types of follicles reveals the important roles of vascular smooth muscle contraction in follicular development in chickens [53]. In Taihe black-bone silky fowls, vascular smooth muscle contraction is deemed as a critical signaling pathway that would affect ovarian development at different egg-laying stages, and ECM-receptor interaction in ovaries might be essential for the transition from laying to broodiness [16, 54]. In pigeons, ECM-receptor interaction and vascular smooth muscle contraction were reported to be closely related to promoting follicular maturation and ovulation in pre-ovulatory follicles [55]. In geese, vascular smooth muscle contraction was involved in follicular development from F4 to F1 stage, and ECM-receptor interaction was significantly enriched in ovaries from the pre-laying period to the broody period [32, 56]. GnRH signaling pathway is known to involve in ovarian function of chickens [57]. In this study, a total of 14 down-regulated DEGs in this study, including CACNA1C, GNAQ, ADCY5, and MMP2, were mapped to GnRH signaling pathway. GNAQ and CACNA1C has been found to be associated with the development of ovarian follicles and onset of the reproductive maturation in animals [58, 59]. In chickens, the expression of GNAQ and CACNA1C were demonstrated to be regulated by MSTRG.19756.2, a novel lncRNA, in trans, and then they acted upstream factors of GnRH signaling pathway to affect the downstream genes involved in ovarian development [60]. ADCY5 as a member of the adenylatecyclases family is reported to be responsible for egg production in Muscovy ducks [37]. Furthermore, ADCY5 has been identified to be closely associated with ovarian morphological related traits of animals [61]. MMP2 is reported to be associated with ovary development of chickens [62]. It was reported that there was lower expression of MMP2 in the atrophy of chicken ovaries [62].
It is now generally recognized that the control of avian ovarian development involves pituitary gonadotropins and various local paracrine and autocrine growth factors such as epidermal growth factor (EGF). In the present study, EGFR, the receptor of EGF, was observed to decrease significantly in ovaries from incubating Changshun hens. Similarly, previous study revealed the dynamic expressions of EGFR in chicken ovarian follicles [63]. Transcriptomic analysis reveals that EGFR is abundantly but differentially expressed in granulosa cells proximal and distal to the germinal disc of chicken preovulatory follicles [64]. EGFR has been shown to be expressed highly in granulosa cells from the prehierarchical follicles, and thereafter its expression is found to decrease markedly to the stage of the largest preovulatory follicles [63, 65]. EGF functions as a ligand of EGFR to stimulate the proliferation of chicken granulosa cells [31, 63, 66]. However, the in vitro experiments have shown that EGF or FSH-induced proliferation of granulosa cell can be reversed by EGFR inhibitor AG1478 [63]. Similarly, AG1478 can also inhibit significantly EGF or FSH-reduced apoptosis of granulosa cells [63]. These results indicate the important roles of EGFR in chicken reproduction and broodiness.
Growth and maturation of ovarian follicles in chickens require a complex network of blood vessels. Compared to prehierarchical follicles which have limited number and size of blood vessels, the large yellow follicles and preovulatory follicles are highly vascularized [49]. The vascular endothelial growth factor (VEGF), platelet-derived growth factor (PDGF), angiopoietin and their receptors are involved in angiogenesis. VEGF is a key regulator of physiological angiogenesis as it can facilitate blood vessel growth and remodeling processes. The network of ovarian blood vessels is closely associated with VEGF. VEGF exerts biological effects by binding to its tyrosine kinase family receptor FLT1, KDR, FLT4 and KDRL (also known as VEGFRKDRL) [67, 68]. Interestingly, VEGF seemly binds to its receptor FLT1 and KDR with high affinity [69]. KDR is mainly expressed in the vascular endothelium of the theca layer in chicken follicles. KDR might be the most important receptor which is involved in VEGF-induced angiogenesis [70–72]. FLT1 can regulate the VEGF activity via interacting with VEGF and making it less available to KDR [73]. Decreased expression of VEGF, FLT1 and KDR might be related to follicle atresia in chickens [49]. Consistently, results from the present study showed that there was a significant decrease in expression of VEGF, including VEGFA and VEGFD, and its receptors, FLT1, KDR and VEGFRKDRL, in ovaries from incubating chickens, indicating the important roles of VEGF and its receptors in chicken broodiness.
Simultaneously, our results showed that there was a significant decrease of PDGF and its receptor (PDGFR) in ovaries of incubating chickens compared with egg-laying hens. There are four PDGF, including PDGFA, PDGFB, PDGFC and PDGFD, and two receptors, including PDGFRA and PDGFRB, identified in vertebrates [74, 75]. The importance of PDGFRA has been confirmed by studies that showed the presence of PDGFRA in follicular cells in the ovaries of different species. In humans, PDGFRA is widely expressed in oocytes, theca cells and ovarian stroma cells [76]. In rats, PDGFRA is also identified in oocytes and granulosa cells [77]. The expression pattern of PDGFRA indicated that it might be involved in the ovarian folliculogenesis, selection and growth initiation of follicles and the formation of thecal layer [78, 79]. Meanwhile, PDGFRA is reported to be a requirement in steroid-producing cells in mice ovaries, and is involved in the steroidogenesis through regulating the downstream target gene Sgpl1, Plekha1, Tiparp, Schip1, and BC058969 [80]. Additionally, signaling downstream of PDGFRA has been reported to induce both apoptotic and antiapoptotic responses [81, 82]. The identification of PDGFRA but not PDGFRB in ovaries from incubating chickens in the present study further suggests that PDGFRA may be important for ovarian development during chicken broodiness.
TEK is a tyrosine kinase receptor and can bind with angiopoietins. The angiopoietin 1 (ANGPT1), angiopoietin 2 (ANGPT2), and TEK receptor tyrosine kinase are mainly expressed in ovaries. In this study, we found TEK and angiopoietins were down-regulated in ovaries from incubating Changshun chickens. The angiopoietin-TEK system is observed to play a crucial role in blood vessel formation and stability, follicular development and atresia [83, 84]. The ANGPT1 and ANGPT2 can bind to TEK, inducing opposite effects. ANGPT1 elicits an activation of TEK by increased tyrosine phosphorylation of TEK when they bind. ANGPT2 as a natural antagonist acts to inhibit the activation of TEK and disrupts ANGPT1-dependent TEK-mediated angiogenesis [84]. ANGPT2 is reported to destabilize existing vessels, loosening the supporting cell matrix to allow angiogenic factors such as VEGF to stimulate cell proliferation and migration during early angiogenesis [85]. Additionally, TEK is hypothesized to play an important role in folliculogenesis [86].
Functionality of the mammalian Kit system, composed of Kit ligand (KL) and its tyrosine kinase receptor (KIT), has been shown that they have multiple roles during oogenesis, folliculogenesis, and melanogenesis [87, 88]. The ovarian expression pattern revealed that mammalian KIT is mainly expressed in theca cells, oocytes and follicular fluid [89–91]. Similarly, the chicken KIT is observed to express in very small follicles (< 1 mm), theca cell layer and the ovarian stroma, indicating that Kit system might promote the transition from quiescence to slow growing follicles in chickens [92]. Furthermore, KIT is reported to be detected in many other chicken organs including the testis, brain, bursa, spleen, thymus, heart and kidney, indicating that Kit system might be implicated in a variety of non-ovarian functions [93].
Fibroblast growth factor receptor 3 (FGFR3) is known to be a receptor for fibroblast growth factors (FGF). In mammals, FGFR3 is identified in oocytes, granulosa cells, theca cells and stromal cells [94, 95]. Additionally, FGFR3 is expressed by human primordial germ cells during the first and second trimester, and is then repressed after meiotic initiation to form primordial oocytes [96]. Study from buffalos revealed that FGFR3 is expressed widely in ovarian follicles during different stages of development [97]. FGFs and their receptors are reported to be involved in multiple biologic processes in angiogenesis, hematopoiesis, wound healing and even embryonic development [98]. Studies based on transgenic mice models showed that mutant FGFR3 leads to dwarfism and infertility [99]. In chickens, FGFR3 is identified to be a candidate gonadal sex differentiation gene in embryo [100]. Decreased FGFR3 in incubating hens in the present study further supports the assumption that FGFR3 might play critical roles in chicken reproduction and broodiness.
In conclusion, we characterized and evaluated the ovarian transcriptome in LP and BP Changshun hens. The results suggest that focal adhesion, MAPK signaling pathway, FoxO signaling pathway, vascular smooth muscle contraction, ECM-receptor interaction and GnRH signaling pathway are critical for incubation behaviour in Changshun hens. Eight candidate genes (EGFR, VEGFRKDRL, FLT1, KDR, PDGFRA, TEK, KIT and FGFR3) in ovaries might play crucial roles in chicken broodiness. Our results provide a theoretical basis for further exploration of the molecular mechanism of broodiness in chickens.
{kind=link}