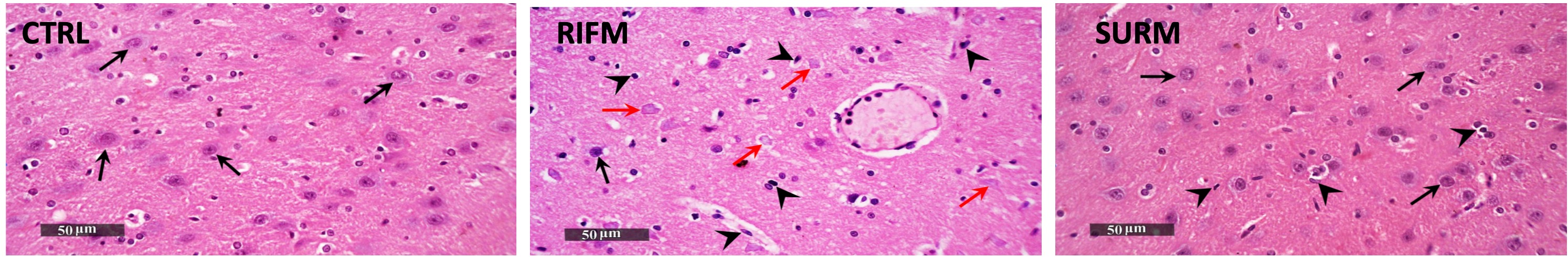
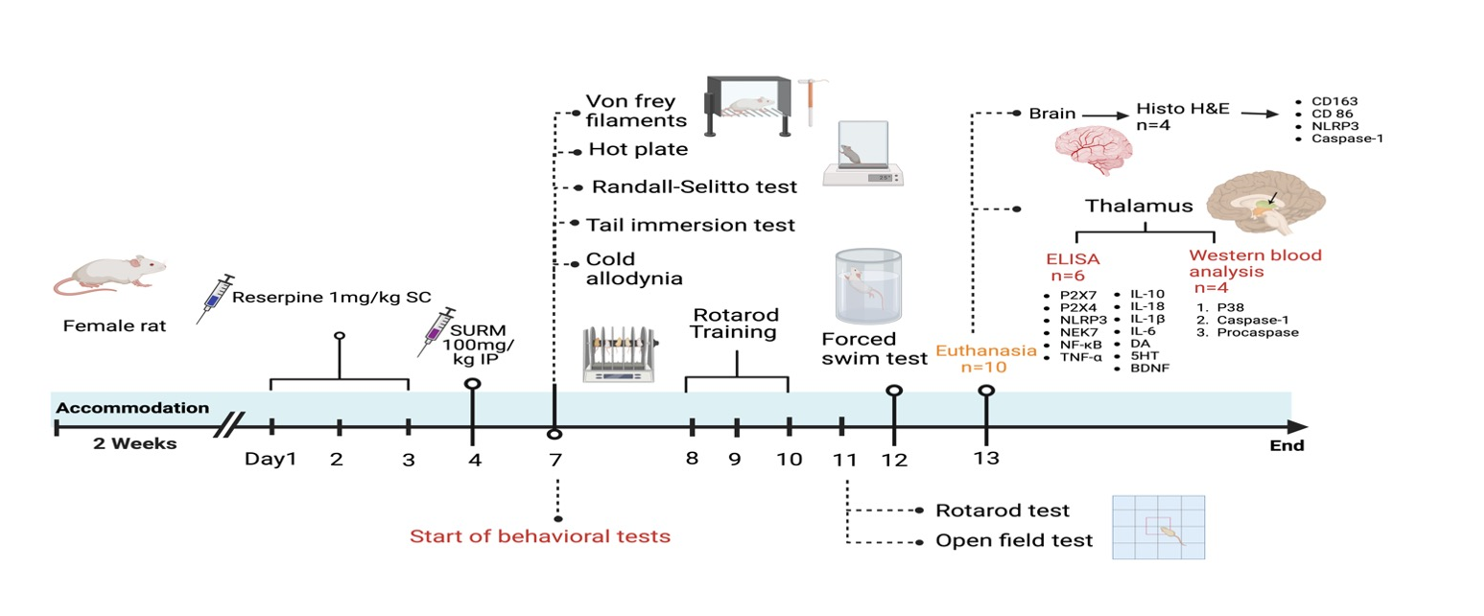
Fibromyalgia syndrome represents a severe manifestation of chronic, widespread pain. A clinical trial demonstrated alterations in thalamic interaction with the cortex, rendering FM brains more sensitive to external stimuli and amplifying pain perception (Kim et al., 2021). Flodin et al. (2014) identified a correlation between elevated connectivity between the left thalamus and prefrontal cortex and heightened pain sensitivity, indicative of hyperalgesia (Flodin et al., 2014). Notably, the data in the present study align with prior research. Histopathological examination of the brains of RES rats using hematoxylin and eosin staining revealed morphological changes in the thalamus compared to healthy rats. Furthermore, this study demonstrated changes in nociception induced by RES injection, evidenced by decreased mechanical allodynia threshold in the von Frey filament test (VFF), decreased cold allodynia paw withdrawal latency (CAPWL), and decreased tail withdrawal latency (TWL) in the tail immersion test. These findings indicate a pronounced allodynia response. Simultaneously, the reduced RSMT and HPWL confirmed a significant hyperalgesia response.
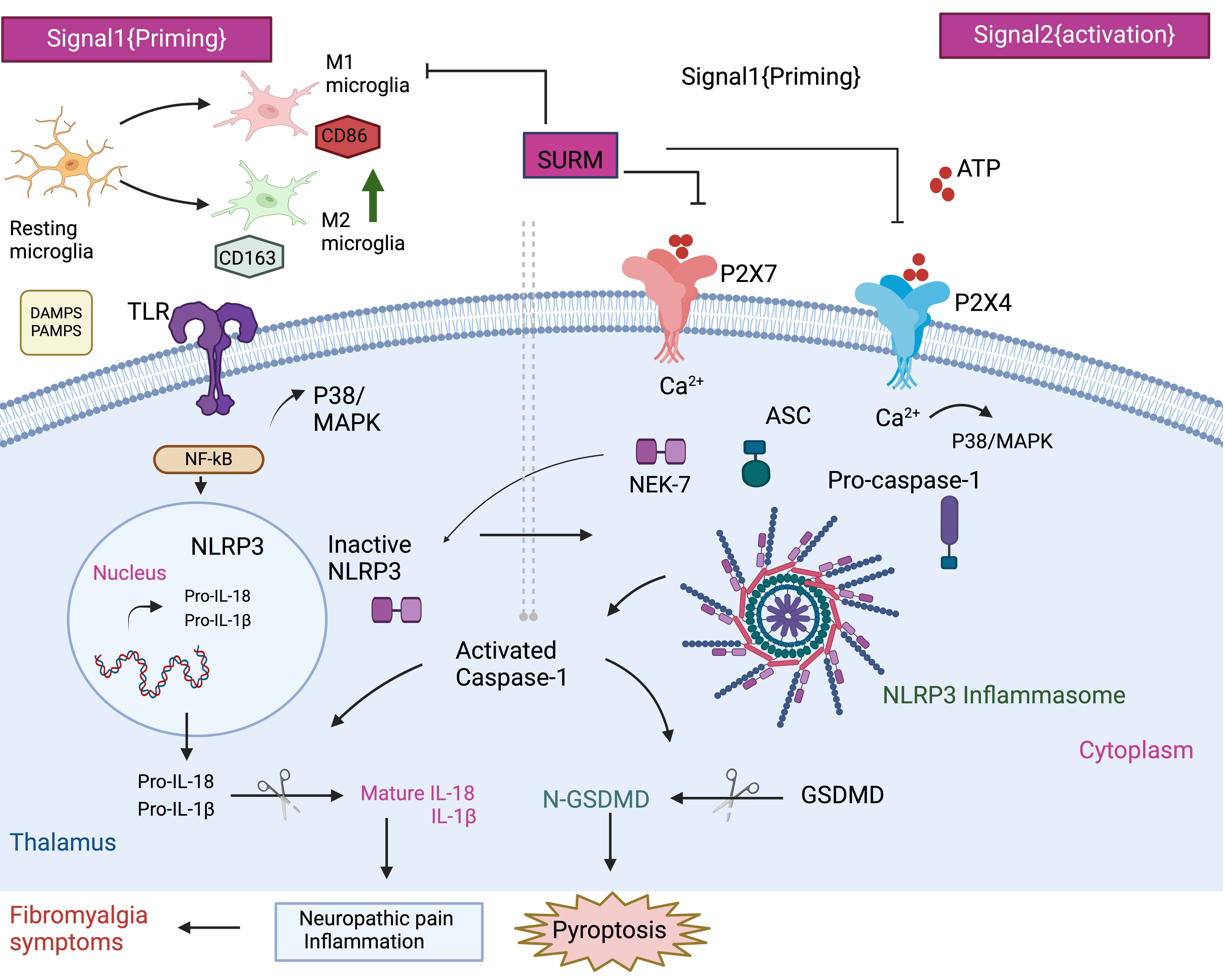
P2X receptors are widely distributed throughout the human body and function as ATP-gated cation channels. These receptors are classified into seven subtypes (1–7), with P2X7 and P2X4 closely associated with the onset of various diseases (Di Virgilio et al., 2017; Stokes et al., 2017). P2X7 is prominently expressed in microglia, playing a significant role in driving neuroinflammation [(Sperlágh and Illes, 2014; Adinolfi et al., 2018). Similarly, P2X4 receptors are abundantly found in neurons and glial cells in several brain regions, including the thalamus (Montilla et al., 2020). Studies have shown that P2X7 and P2X4 receptors are closely linked to pain mechanisms [(Ulmann, Hirbec and Rassendren, 2010; Hu et al., 2022). Interestingly, in the current study, inhibition of P2X7 and P2X4 receptors by SURM alleviated hyperalgesia in the Randall-Selitto and hot plate tests, as well as allodynia in the VFF, cold allodynia, and tail immersion tests. This demonstrated significant anti-hyperalgesic and anti-allodynic responses to purinergic receptor inhibition, highlighting the pivotal role of purinergic receptors in pain transmission through the thalamic tract in fibromyalgia. The co-expression of P2X4 and P2X7 receptors enhances the P2X7 receptor-mediated inflammatory response and facilitates P2X7 receptor-mediated IL-1β production and inflammation through calcium influx (Kawano et al., 2012). Activation of P2X7 is critical in the assembly of the NLRP3 inflammasome and subsequent release of pro-inflammatory cytokines like IL-1β and IL-18 [(Di Virgilio et al., 2017). Importantly, this process requires calcium influx triggered by the P2X4 receptor (Sakaki et al., 2013), illustrating the neuroprotective effect of purinergic receptor blockade against the release of pro-inflammatory cytokines.
Previous research has indicated the critical role of the NLRP3 inflammasome in pain plasticity and its involvement in inflammatory disorders (Mao et al., 2020). The NLRP3 inflammasome is a crucial signaling hub regulating the production of IL-1β and IL-18, two pro-inflammatory cytokines of the IL-1 family (Broz and Dixit, 2016; Jo et al., 2016). Inflammasome activation is regulated by two signals: the Toll-like receptor (TLR)/nuclear factor (NF)-κB pathway as the first signal and the second signal involving damage-associated molecular patterns (DAMPs) and pathogen-associated molecular patterns (PAMPs). These signals converge to form a multi-protein complex comprising pro-caspase-1, NLRP3, and the adaptor protein ASC, activating the functional NLRP3 inflammasome (Hornung and Latz, 2010). Upon assembly of the NLRP3 inflammasome, inactive caspase-1 is cleaved into its active form by proteases. In the present study, ELISA and tissue-based immunohistochemical analyses demonstrated reduced immunoreactivity for both NLRP3 and active caspase-1 in the thalamus of SURM-treated rats compared to untreated animals. This reduction may be attributed to inhibition of the TLR/NF-κB pathway or P2X7/P2X4 receptors. NEK7, a family member of the NIMA-related kinases (NRKs), regulates NLRP3 activity (Fry et al., 2012). SURM inhibits P2X7, which controls NEK7, potentially contributing to the reduced NLRP3 expression. Notably, the P2X4 receptor regulates NLRP3 protein production in a diabetic nephropathy model (Chen et al., 2013), and its activation can activate NF-κB upstream of NLRP3 (Xing et al., 2023). NLRP3 gene expression may be up-regulated in FM rats due to NF-κB or P2X4, whereas SURM administration blocks this effect (Shen et al., 2023).
Earlier research has investigated the NF-κB/NLRP3 inflammasome signaling pathway as a specific target for managing neuropathic pain (Zhang et al., 2017). This idea stems from previous findings indicating that NF-κB signaling plays a crucial role in nociception in chronic pain disorders like fibromyalgia (Kaur et al., 2019) and other chronic pain conditions (Lee et al., 2004; Luo et al., 2020). Immunohistochemical studies on fibromyalgia muscles have shown strong expression of NF-κB, with IL-1β and IL-18 being NF-κB-dependent pro-inflammatory cytokines (Cordero et al., 2014). Using the RIFM rat model, it becomes possible to demonstrate a positive correlation between NF-κB and NLRP3 inflammasome activation in promoting the production of inflammatory cytokines, leading to hyperalgesia. Interestingly, SURM-treated rats exhibited inhibition of NF-κB and subsequent inflammasome activation. This inflammatory cascade contributes to resting macrophage transition towards M1 activation over M2, leading to neuroinflammation (Sui et al., 2020). In various neuropathic pain models, high expression of inflammatory M1 macrophage-specific molecules and low levels of M2 microglia-specific molecules indicate M1/M2 microglia polarization shift (Kiguchi et al., 2015, 2017; Atta et al., 2023a). In fibromyalgia patients, nociplastic pain arises due to M1 microglia-dependent neuroinflammation [(Tripathi et al., 2021; Atta et al., 2023b). This aligns with the current study's findings, as immunohistological results in RIFM rats showed a shift towards the M1 phenotype, as seen in CD163, a marker of M1 microglia. Remarkably, after inhibiting NF-κB signaling with SURM, M1-polarized microglia decreased, and M2-polarized microglia increased in the thalamus of RIFM rats. This reversal in microglia polarization aligns with the highly dynamic nature of this process observed in both healthy and pathological conditions. Additionally, this aligns with Suramin's anti-inflammatory benefits observed in vitro in intervertebral disc degeneration, partly linked to its interference with NF-κB signaling (Liu et al., 2021).
Caspase-1 is critical in pyroptosis, an inflammatory form of cell death (Shi et al., 2015). Pyroptosis involves gasdermin D (GSDMD) and interleukins such as IL-1β and IL-18. Active caspase-1 converts pro-IL-1β and pro-IL-18 into their biologically active forms (Davis, Wen and Ting, 2011). Additionally, active caspase-1 cleaves GSDMD, creating pores in the plasma membrane that enable the release of IL-1β and IL-18. In the current study, RIFM rats exhibited significantly elevated levels of GSDMD compared to normal animals. This increase may be attributed to P2X7 and P2X4 receptors. This aligns with previous findings showing that silencing P2X7 alleviates pyroptosis-associated conditions like abdominal aortic aneurysm (Sun et al., 2023). Mechanistically, silencing P2X4 blocks the NLRP1/caspase-1 pathway, demonstrating a neuroprotective effect in an intracerebral hemorrhage mouse model, while administration of an NLRP3 activator induces neuronal impairment in the same model (Wu et al., 2023). This indicates a strong relationship between P2X4 and the NLRP3 inflammasome. Furthermore, activation of the P2X7 receptor leads to MAP kinase (MAPK) activation downstream (Suzuki et al., 2004). Studies on thalamic injury (Deng et al., 2023) and peripheral nerve injury (Terayama et al., 2008) have highlighted the involvement of microglial p38-MAPK in neuropathic pain development. SURM-treated rats showed a significant reduction in p38 expression compared to RIFM rats. This may explain SURM's behavioral effects, as neuropathic pain and central sensitization are mediated by the activation of P2X7/P2X4 receptors and p38-MAPK (Ji and Suter, 2007).
IL-1β and IL-18 are involved in neuroinflammation and pain processes peripherally and centrally (Verri et al., 2007; Ren and Torres, 2009). Notably, serum samples from FM patients have shown elevated levels of IL-1β and IL-18. Furthermore, IL-1β serum levels correlate positively with FM patients' pain scale scores(Cordero et al., 2014). The current study aligns with these previous findings, as SURM administration reduced the expression of IL-1β and IL-18 in the thalamus compared to RIFM rats. This aligns with research showing that inhibiting the thalamic NLRP3 inflammasome reduces IL-18 and IL‑1β, leading to behavioral alleviation of thermal and mechanical pain sensitivity in a central post-stroke pain mouse model (Huang et al., 2022). The pro-inflammatory cytokine IL-1β may affect BDNF expression through P2X4 receptor activation on microglia (Guo et al., 2020). This may attenuate BDNF's neuroprotective effects by IL-1β-mediated MAPK activation, as observed in in vitro studies (Tong et al., 2008). In the current study, RIFM rats exhibited decreased BDNF expression, while SURM-treated rats showed a reversal of this decrease, indicating a potential neuroprotective effect of SURM.
Additionally, the delayed production of IL-10 has been implicated in promoting hyperalgesia (Okamoto et al., 2001). Nerve injury often leads to an imbalance between pro-inflammatory cytokines such as IL-1β, IL-18, IL-6, and TNF-α and anti-inflammatory cytokines like IL-10 (Alcami, 2003). In FM rats, elevated levels of TNF-α and IL-6 in the thalamus may contribute to hyperalgesia. This heightened inflammatory state could explain depressive behavior, similar to observations in post-mortem studies of depressed suicide victims (Steiner et al., 2008; Suzuki et al., 2019). Previous studies have linked elevated allodynia and hyperalgesia in FM rats to depressive-like behavior (Blasco-Serra et al., 2015). However, treatment with SURM reduced these cytokine levels, indicating an anti-inflammatory response through increased IL-10 production. SURM-treated rats showed reduced immobility time in the forced swim test (FST), suggesting an anti-depressive effect. This could be attributed to the depression of pro-inflammatory cytokines or P2X7 inhibition, as seen with the blockade of P2X7 receptors using brilliant blue G, which reduced depressive-like behavior in RIFM rats (D’Amico et al., 2021b). Furthermore, treatment with SURM restored catecholamine levels in the thalamus of rats. This restoration may be linked to NLRP3 inhibition, as observed in studies where stem cell treatment restored brain monoamines in FM models (Mokhemer et al., 2023).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}