Decidualization is a tissue specific differentiation of endometrial stromal cells into epithelial-like secretory cells. From an immunological perspective, it is a multistep process that starts with an acute pro-inflammatory response that allows endometrial receptivity and embryo invasion 30. The duration of this period is tightly controlled, lasting between 2 and 4 days 10,31. The subsequent switch to an antiinflammatory response at the closure of the implantation window is mediated by the emergence of mature decidual cells and the recruitment and activation of uterine NK cells to clear senescent cells which constrain inflammation. This step sustains the transition of the decidua to a semi-permanent tissue critical to support the ongoing pregnancy and the maternal tolerance to the fetus 32,33. However, in the absence of an implanting embryo, the levels of progesterone falling down and the propagation of senescence through the endometrium are associated with influx of neutrophils and macrophages, resulting in tissue breakdown and menstruation 34,35.
The inflammatory response characteristic of the implantation window has been linked to the ER stress, UPR response and senescence that EnSCs undergo during decidualization 8,10,18,20. Here, by means of in silico and in vitro designs, we point to the role of ER stress and UPR in controlling the emergence of senescent cells at decidualization.
First, we used an in silico approach to evaluate the kinetic changes of ER stress and UPR-associated genes through the decidualization timecourse. This analysis highlighted HSPA5, ATF6, ATF4 and TXNIP as abundantly expressed across the process. Moreover, network clustering suggested a specific role for these genes upon decidualization of EnSC during the periimplantation period. In this sense, HSPA5 upregulation occurring during decidual differentiation suggests the participation of BiP chaperone to sustain the dramatic increase of protein secretion upon decidualization. Additionally, a predominant expression of TXNIP in senescent decidual cells might be associated with the activation of the NLRP3 inflammasome, triggering IL-1β release. In contrast, ATF6 rise in mature decidual cells subset might account for its key role in the implantation process. In agreement, pharmacological inhibition of ATF6 pathway prevented trophoblast expansion in an in vitro model of implantation 36. Also, ATF6 levels were found highly expressed in the murine uterus near to the implantation site 37.
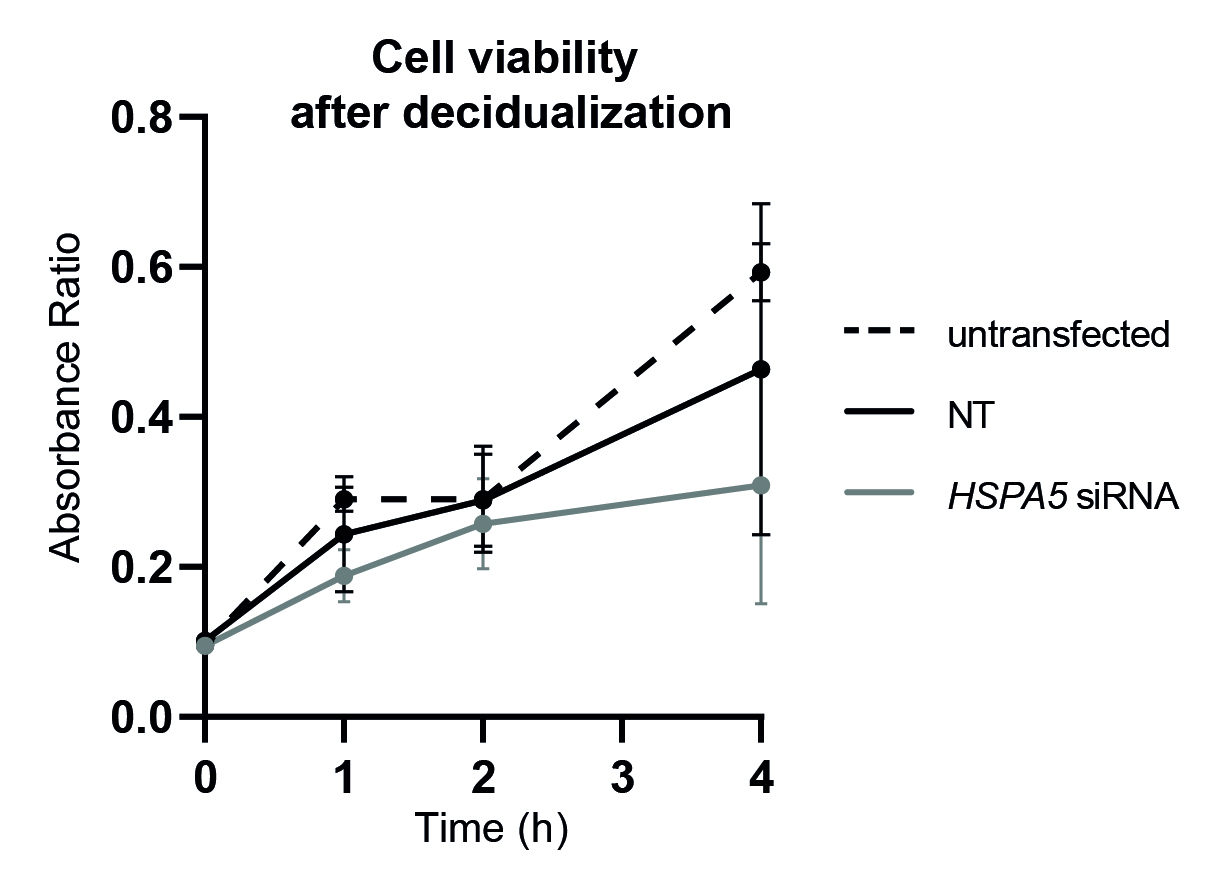
The divergence of EnSC phenotypes into DC and snDC subpopulations was previously characterized during the decidualization process 18,20. On this basis, we focused on the endoplasmic reticulum chaperone BiP, HSPA5, a key regulator of the UPR response. Using an in silico approach, HSPA5 showed a peak of expression just before the divergence, whereas the knockdown of HSPA5 in EnSCs impaired the decidualization program. This was evidenced by the downregulation of two canonical differentiation markers, PRL and IGFBP1. Upon further analysis, we observed that the emergence of both mature and senescent cells was altered. This was confirmed by a lower IL1RL1 expression, as well as a reduced β-galactosidase activity, which are markers of each subpopulation, respectively. It is worth mentioning that, even though the significant reduction of HSPA5 mRNA levels occurred just until day 4, downstream effects of silencing were observed mostly at day 8 of the differentiation process. This observation reinforces the idea of HSPA5 as a biphasic gene, with an early role in the control of the final cell fate of endometrial cells at decidualization.
Adding to the relevance of ER chaperones in the decidualization process, previous reports showed that HSPA8 might play a role in embryo-quality biosensoring 10. Moreover, knockdown of HSPA8 in decidual cells generates an exacerbated UPR, which also compromises the production of PRL and IGFBP1. Here, the knockdown of HSPA5 impaired the progression of the decidual pathway and the divergence of cells into DC and snDC subpopulations.
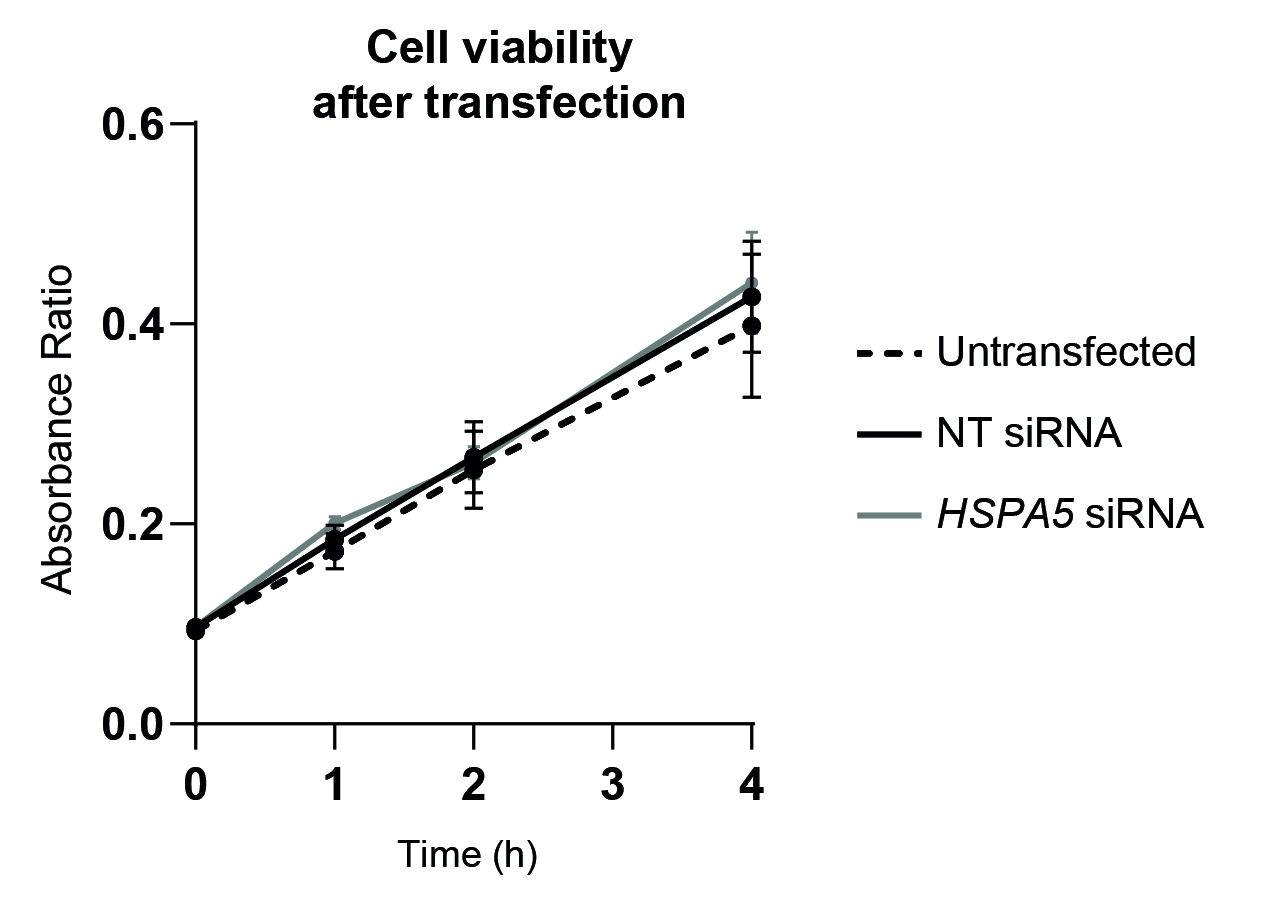
ER stress is triggered upon the accumulation of misfolded proteins inside the endoplasmic reticulum. In the context of decidualization, this is caused by the dramatic increase of secreted factors. Alongside the effect of HSPA5 knockdown on the decidual markers, we observed the induction of higher levels of ER stress sensor proteins IRE1α and PERK. In the light of these observations, it should be considered whether the impaired decidualization is caused by the absence of BiP itself, or if these effects are due to a stronger UPR response in decidualizing cells. While both scenarios are possible and compatible, it is noteworthy that ER stress levels caused by HSPA5 knockdown seemed to be moderate, since cell death was not induced.
The role of BiP in cellular senescence is unclear since its levels were found to be increased 38,39 or decreased 40–44 depending on the cellular context. Moreover, two non-small cell lung cancer models displayed a different effect on HSPA5 expression in response to cisplatin induced senescence 45. Interestingly, in that work Ei et al showed that cells that upregulated HSPA5 expression during senescence induction were able to reenter the cell cycle after cisplatin removal, suggesting a role for BiP in senescence reversal. This observation is particularly interesting in the context of the decidualization process, since the endometrium is a highly dynamic tissue that regenerates itself in each menstrual cycle.
Considering the broad ER stress and UPR pathway, several lines of evidence point to a link with cellular senescence; however, the literature is controversial. A positive association between both processes was reported by Matos et al using a human diploid fibroblasts model. They observed that ER stress was induced not only by replicative senescence, but also by stress-induced premature senescence. Moreover, senescence induced IRE1α and PERK activation, while the inhibition of these pathways prevented senescence features such as β-galactosidase activity and p21 expression 46. In line with this, X radiationinduced senescence also triggered ER stress in pulmonary artery endothelial cells 47. On the contrary, Zhu et al showed a reduction of ER stress levels under oncogene-induced senescence on mouse keratinocytes; while ER stress induction augmented cell proliferation and attenuated βgalactosidase activity 48. This opposite association between ER stress and senescence is in line with the results reported here for decidualizing cells, though deeper studies of the molecular mechanisms and specific UPR pathways involved are needed.
From a clinical perspective, alterations in both ER stress and senescence have been associated with alterations in endometrial receptivity and early pregnancy. In this sense, inhibition of IRE1α endonuclease activity in decidualized cells reduced the ability of trophoblast cells to expand over them in an in vitro model 8. This enzymatic activity controls the cytosolic splicing of XBP1 mRNA, which is a key feature of the UPR process. Interestingly, endometrial samples from patients with Recurrent Implantation Failure show lower levels of the spliced XBP1 mRNA compared to fertile women, suggesting that mild UPR levels are needed for a proper endometrial receptivity 8. On the other hand, patients with Recurrent Pregnancy Loss (RPL) display an aberrant decidual response as well as uNK cell deficiency, which might be associated to impaired clearance of senescent cells 18. The accumulation of senescent cells promotes a persistent inflammation via continuous production of the SASP, socalled bystander senescence, which underpins a pathological process with a gradual loss of organ function. In addition, biosensing of soluble signals from low fitness embryos inhibits uNK cell-dependent removal of senescent decidual cells 26.
The compromised balance between decidualized and senescent cells, as well as altered ER stress levels, might impact endometrial function. In this sense, HSPA5 knockdown impaired colonyforming unit activity of primary EnSC, indicative of loss of cellular plasticity. Interestingly, it has been shown that RPL is strongly associated with uterine stem cell deficiency, subsequently limiting differentiation potential, disturbing decidualization and leading to consecutive miscarriages 49. Moreover, decreased decidual senescence in the context of a clinical trial was associated to upregulated clonogenic capacity of EnSCs 50.
Together, our results show a key role for HSPA5/BiP in reprogramming EnSC and highlight the importance of constraining ER stress levels during this process. Considering the relevance of ER stress and senescence in endometrial homeostasis, as well as their role in fertility complications, the identification of HSPA5 in the control of the decidual pathway progression sets this chaperone as a potential key regulator of the endometrial function. Further studies are needed to elucidate its role in vivo and its prognostic value for endometrial receptivity.
{kind=link}
{kind=link}