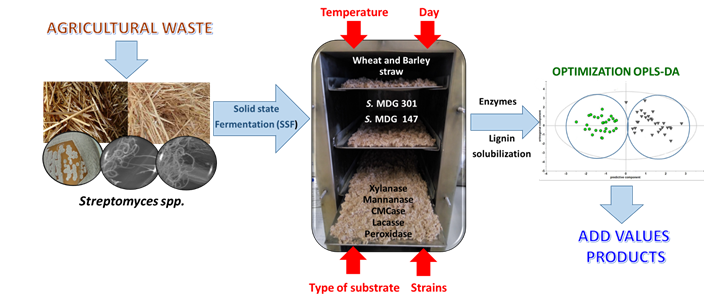
3.1. Multivariate Analysis of the enzymatic activities, APPL and alkali-lignin yields under different SFF conditions.
The degradation of lignocellulose was studied by measuring the activity of three hydrolytic enzyme (xylanase EC 3.2.1.8, mannanase EC 3.2.1.78, and CMCase EC 3.2.1.203) and two oxidative enzyme (laccase EC 1.10.3.2 and peroxidase EC 1.11.1.7). This was done to determine the effect of the fermentation time, type of substrate, temperature and microbial strain on the SSF process in terms of enzymatic activity. Additionally, the content of APPL and alkali-lignin in the extracts obtained from SSF cultures was determined to evaluate the degradation capacity of the two strains on the residues.
Table 1 shows the results of the enzymes analysis on the substrates under the different experimental conditions Also, the values obtained for APPL and alkali-lignin contents in the extracts obtained from fermented residues are summarized in Table 2.
|
Table 1
Extracellular enzymatic activities obtained from wheat and barley straw after SSF process under the different operational conditions tested. All values are given as mean ± standard deviation (n=3).
|
|
Substrate
|
T (ºC)
|
Strain
|
Days
|
Xylanase
(U/mL)
|
CMCase (U/mL)
|
Mannanase (U/mL)
|
|
Wheat
|
28
|
301
|
2
|
0.00±0.00
|
0.00±0.00
|
0.00±0.00
|
|
Wheat
|
28
|
301
|
4
|
0.63±0.06
|
0.01±0.01
|
0.01±0.02
|
|
Wheat
|
28
|
301
|
7
|
0.26±0.10
|
0.03±0.01
|
0.00±0.0
|
|
Wheat
|
28
|
147
|
2
|
1.35±0.29
|
0.90±0.10
|
1.23±0.65
|
|
Wheat
|
28
|
147
|
4
|
8.16±0.20
|
1.76±0.01
|
6.31±0.13
|
|
Wheat
|
28
|
147
|
7
|
6.66±0.34
|
0.88±0.02
|
3.68±0.14
|
|
Wheat
|
45
|
301
|
2
|
31.11±1.21
|
10.19±1.29
|
0.00±0.00
|
|
Wheat
|
45
|
301
|
4
|
32.78±0.16
|
8.76±0.10
|
0.00±0.00
|
|
Wheat
|
45
|
301
|
7
|
33.91±0.52
|
7.43±0.31
|
0.00±0.00
|
|
Wheat
|
45
|
147
|
2
|
0.52±0.05
|
0.15±0.01
|
0.08±0.00
|
|
Wheat
|
45
|
147
|
4
|
4.25±1.75
|
1.18±0.02
|
4.30±0.13
|
|
Wheat
|
45
|
147
|
7
|
1.10±0.51
|
0.00±0.00
|
0.00±0.00
|
|
Barley
|
28
|
301
|
2
|
3.73±1.34
|
0.48±0.05
|
0.72±0.10
|
|
Barley
|
28
|
301
|
4
|
7.15±1.46
|
0.61±0.13
|
0.28±0.23
|
|
Barley
|
28
|
301
|
7
|
7.47±1.52
|
0.53±0.12
|
0.04±0.04
|
|
Barley
|
28
|
147
|
2
|
1.50±0.56
|
0.59±0.23
|
0.00±0.00
|
|
Barley
|
28
|
147
|
4
|
1.07±0.08
|
0.00±0.00
|
0.00±0.00
|
|
Barley
|
28
|
147
|
7
|
0.87±0.01
|
0.34±0.16
|
1.16±0.10
|
|
Barley
|
45
|
301
|
2
|
5.35±0.40
|
1.25±0.05
|
1.22±0.11
|
|
Barley
|
45
|
301
|
4
|
22.85±1.88
|
1.16±0.09
|
0.77±0.12
|
|
Barley
|
45
|
301
|
7
|
27.95±1.58
|
1.55±0.2
|
0.69±0.19
|
|
Barley
|
45
|
147
|
2
|
1.13±0.63
|
0.46±0.070
|
0.00±0.00
|
|
Barley
|
45
|
147
|
4
|
2.433±0.61
|
0.17±0.00
|
0.37±0.14
|
|
Barley
|
45
|
147
|
7
|
5.71±1.35
|
0.30±0.04
|
1.15±0.01
|
|
Table 2
APPL and alkali-lignin yields obtained from the wheat and barley straws after the SSF process under the different tested operational conditions. All values are given as mean ± standard deviation (n=3).
|
|
Substrate
|
T (ºC)
|
Strain
|
Days
|
APPL (g)
|
Alkali-lignin (g)
|
|
Wheat
|
28
|
301
|
2
|
0.10±0.01
|
0.21±0.01
|
|
Wheat
|
28
|
301
|
4
|
0.14±0.03
|
0.24±0.01
|
|
Wheat
|
28
|
301
|
7
|
0.17±0.00
|
0.61±0.00
|
|
Wheat
|
28
|
147
|
2
|
0.03±0.01
|
0.55±0.03
|
|
Wheat
|
28
|
147
|
4
|
0.10±0.00
|
1.31±0.02
|
|
Wheat
|
28
|
147
|
7
|
0.06±0.01
|
1.46±0.02
|
|
Wheat
|
45
|
301
|
2
|
0.17±0.00
|
0.54±0.01
|
|
Wheat
|
45
|
301
|
4
|
0.11±0.00
|
0.72±0.01
|
|
Wheat
|
45
|
301
|
7
|
0.17±0.00
|
1.38±0.01
|
|
Wheat
|
45
|
147
|
2
|
0.07±0.03
|
0.49±0.01
|
|
Wheat
|
45
|
147
|
4
|
0.10±0.00
|
0.43±0.02
|
|
Wheat
|
45
|
147
|
7
|
0.11±0.00
|
1.35±0.12
|
|
Barley
|
28
|
301
|
2
|
0.09±0.01
|
0.62±0.19
|
|
Barley
|
28
|
301
|
4
|
0.06±0.00
|
0.57±0.14
|
|
Barley
|
28
|
301
|
7
|
0.09±0.00
|
0.49±0.01
|
|
Barley
|
28
|
147
|
2
|
0.05±0.00
|
0.54±0.00
|
|
Barley
|
28
|
147
|
4
|
0.03±0.01
|
0.50±0.02
|
|
Barley
|
28
|
147
|
7
|
0.04±0.00
|
0.50±0.00
|
|
Barley
|
45
|
301
|
2
|
0.07±0.02
|
0.35±0.03
|
|
Barley
|
45
|
301
|
4
|
0.08±0.01
|
0.43±0.06
|
|
Barley
|
45
|
301
|
7
|
0.13±0.01
|
0.46±0.05
|
|
Barley
|
45
|
147
|
2
|
0.07±0.02
|
0.53±0.01
|
|
Barley
|
45
|
147
|
4
|
0.07±0.02
|
0.47±0.02
|
|
Barley
|
45
|
147
|
7
|
0.09±0.01
|
0.45±0.02
|
In all the extracts obtained from both fermented substrates, neither laccase nor peroxidase activities were detected, probably due to the adsorption of these enzymes to the substrate, which makes their recovery difficult [11].
These data were analysed using OPLS-DA, evaluating the individual effect and the interaction of the variables studied in terms of the enzymatic response.
The OPLS-DA model indicated that the discrimination among the Streptomyces strains and substrates-type levels was already clear after calculating three predictive t(1-3) components. In other words, the fermentation using wheat and barley straws, combined with any of these strains showed a clearly differentiated pattern. The R2X(1) parameter indicated that about 24.1 % of the X main-data matrix variation was modelled by the t [1] predictive component. Likewise, R2X(2) and R2X(3) showed that 20.5 % and 11.5 % of the X main-data matrix variation was modelled by the t(2) and t(3) predictive components, respectively. Moreover, a total 25 % of the X variation was modelled by three to(1-3) orthogonal components. Thus, a total 81.1 % of the X variation was modelled by all the (predictive and orthogonal) components, which renders a rather good explanation of the separation. The outliers outside the 95 % confidence Hotelling’s T2 ellipse were not excluded from the analysis because, in perspective, they did not affect the overall separation results.
Figure 1 shows the 3D Scores Scatter plot from the OPLS-DA model used to test the effect of the substrate type and the strains employed in the SSF process for the enzymatic production. In both cases, it can be observed that it is possible to clearly discriminate the effect of each studied variable.
On the other hand, despite that the incubation temperature showed a less homogeneous groups distribution, it allowed differentiating two distinct effects. Additionally, the incubation time did not show a homogeneous behaviour, possibly because this parameter is a determining factor of the studied variables (Figure 2).
3.2. Optimized enzyme activities in the bio-treated substrates
According to the OPLS-DA model, the level of xylanase production during the incubation time showed clear differences between the two evaluated Streptomyces strains. Thus, the combined observation of the model showed in Figure 1, 2 and 3a, indicates that the maximum xylanase activities were obtained in fermented wheat straw with S. MDG 301 strain, after 7 days of incubation at 45 °C (dot 1). It is important to highlight that no apparent differences were observed between the maximum xylanase production in wheat straw at the above-mentioned conditions and the xylanase production after 2 and 4 days (Figure 3a, coloured region). Therefore, the obtained data suggest that high production of this enzyme can be reached in shorter fermentation periods, which implies an important advantage for industrial applications due to the consequent reduction in production time and cost. Similarly, in fermented barley straw, despite obtaining a general lower enzyme production in comparison to wheat, the results showed that a few samples had a significant xylanase production, in the same conditions (Figure 1, 2 and 3a, dot 2). This aspect was not evidenced by ANOVA analysis which revealed large significant differences (Pr > F: 0.0006) between the wheat (Figure 1, 2 and 3a, dot 1) and the barley (Figure 1, 2 and 3a, dot 2) samples.
The high levels of xylanase obtained in both fermented substrates are a consequence of their chemical composition. Agricultural residues, including wheat and barley straw, contain high amounts (approximately 25 % w/w) of hemicellulosic sugars, mainly xylose, which induces the production of this enzyme [41].
Regarding the CMCase activity, the OPLS-DA analysis showed a higher production rate of this enzyme in the wheat straw after 2 days of incubation with the S. MDG 301 strain at 45 °C (Figure 1, 2 and 3b, dot 3). In the barley straw, the differences found throughout the incubation time were not significant and the enzyme production was significantly lower than those obtained in wheat straw. However, low concentrations of mannanase enzyme activity were detected in these conditions. This result could be attributed to the practically absence of inducers of this activity in the residues, corresponding the activity detected to the enzyme basal or constitutive level. On the contrary to the conditions obtained for the maximum production of xylanase and CMCase activities, the highest activity of the mannanase, was detected using wheat straw treated with the S. MDG 147 strain incubated at 28 °C during 4 days (Figure 1, 2 and 3c, dot 4).
3.3. Optimized APPL and alkali-lignin yields from fermented substrates
According to the OPLS-DA model the amount of solubilized APPL was significantly higher using wheat straw as substrate, and when it was inoculated with the S. MDG 301 strain. The maximum solubilisation of APPL was reached on the day 7 (Figure 1, 2 and 4a, dot 1). In order to increase the extraction yield of solubilized lignin from the fermented substrates, an alkaline extraction with 0.1 M NaOH was carried out. As expected, the amount of lignin solubilized from the wheat and barley straws using alkali was higher than using water. Thus, as estimated, these results coincided with the highest enzymatic activities which could be attributed to the fact that the enzymatic cleavage of the structure bonds is bound to the consequent substrate solubilisation [34]. It is important to take into consideration that the reported arabinoxylan/glucomannane ratio in non-aqueous angiosperms is 20:1 [11]. For this reason, the main role of the APPL solubilisation should be mostly ascribable to the production of xylanase activity for the chemical composition of the substrate [42].
The highest lignin solubilization (APPL) and highest alkali-lignin extractions were obtained from wheat straw fermented with the S. MDG 147 strain, after 7 days of fermentation at a temperature of 28 ºC (Figure 1, 2 and 4b, dot 5). The synergistic action of xylanases, mannanases and CMCase activities produced by this strain might facilitate the extraction of lignin by the alkali. Similar results were obtained from wheat straw fermented with S. MDG 301 strain after 7 days of incubation likely due to the enhanced solubilization caused by enzymatic action of xylanase (Figure 4a, dot 1).
Undoubtedly, lignocellulosic biomass is considered a low cost and easy to acquire residue that can be revalorized and used as fermentable substrate to produce inputs/raw materials of industrial interest. The use of products generated by SSF of lignocellulosic residues have increased significantly in the pharmaceutical, food, lubricants, paper, and textile industries, acknowledging the potential of depolymerizing enzymes to generate favourable chemical changes in substrates [43-45]. Hence, among the parameters influencing the fermentation process, the enzymes production is the most relevant aspect. However, it is important to emphasize that the economy, efficiency and success of such biotechnological processes largely depend on the control of other crucial aspects, including the substrate nature, the microorganisms employed, and the process operational conditions [46]. These parameters will in turn determine the enzymatic activity reached, the extent of the chemical changes in the substrates, and thus, the production of bio-products with specific characteristics.
For instance, we previously reported that lignocellulosic residues, microbiologically treated and subsequently functionalized, were successfully used as thickeners to elaborate lubricant greases. The rheological and tribological characteristics of the new greases largely depended on the substrate employed and the activity of the enzymes involved in the lignin depolymerization process. A clear increase in the rheological and viscoelastic properties were directly related to the reduction of the soluble lignin. This is due to the β-O-4 bonds disruption, together with an increase in the C, H, and N content due to the influence of the xylanase enzyme activity [34]. However, in specific cases, the desired substrate changes occur with a minimal enzyme production. For instance, Rajeev et al., (2009) [47] studied the ethanol production by saccharificating three different feedstocks (rice straw, sugarcane bagasse and water hyacinth biomass). They showed that the highest reaction yield occurred in SSF with rice straw, whose saccharification process occurred with low cellulase activity. In these studies, the production of the desired metabolite was clearly governed by the paramount enzymatic action, which in turn was dependent on the optimization of the specific process control variables.
Multivariate analysis can concurrently optimize the experimental variables in order to find the best operational conditions for biotechnological processes and discriminate systems regarding one or more interesting features. At the same time, it helps determining the value of multiple physical-chemical properties directly or indirectly related to the substrate or product chemical composition. Therefore, the use of conventional biochemical analyses and powerful biostatistical tools (OPLS-DA model) represents a natural marriage. It allows to determine the most suitable conditions for maximising the production of a metabolite of industrial interest (e.g., specific enzymatic production), considering the overall interaction of the operating variables that affect such processes. This integrated approach provides valuable information to set up the appropriate fermentation conditions to produce the desired chemical changes in the substrates, and thus, to get a particular product of industrial interest.
{kind=link}