The vocal fold (VF), an essential apparatus for phonation, vibrates hundreds of times a second.1 Due to its role and anatomical location, the VF is inherently exposed to mechanical and environmental stress. As many as 20 million people report voice disorders annually in the US.2 Inflammation is broadly associated with dysphonia and is likely etiologic for benign vocal fold lesion development. Glucocorticoids (GCs) are frequently administered to manage voice disorders because of their anti-inflammatory functions and affordability.3–6 However, recent reports suggest clinical outcomes of GC therapy to be variable.7–9 Diverse functions of GCs likely underlie the disparate outcomes.10–12 Of note, fibrotic effects of GC signaling have been reported in several organs and VF fibroblasts.13–15 Optimizing GC therapy to minimize fibrosis while limiting inflammation has the potential to benefit millions of patients.
In the inflammatory milieu, dysfunctional cooperation between tissue-resident and infiltrated hematopoietic cells can drive pathological tissue responses, such as chronic inflammation and fibrosis.16 Previous studies revealed the significance of intercellular communication between fibroblasts and macrophages in pathologies across multiple organs, including the VF.17–20 Differential macrophage phenotypes are induced via exposure to stimuli through the shift from inflammatory to fibrotic environments.17,21,22 Inflammatory stimuli, such as interferon-gamma (IFN- γ) and lipopolysaccharide (LPS), induce the inflammatory M1 phenotype.17,23 Interleukin (IL)-4, IL10, and transforming growth factor-β (TGF-β) drive the anti-inflammatory/fibrotic M2 phenotype.23–25 However, various subtypes beyond the dualistic classification to the M1 and M2 are induced by individual stimuli,17 likely related to organ-specific responses to macrophages.26,27 For example, in VF fibroblasts, fibrotic genes were not activated by paracrine signaling from IL4-stimulated typical M2 macrophages,27 which elicited a fibrotic response in non-VF fibroblasts.28,29 Conversely, TGF-β-stimulated macrophages induced a fibrotic response in VF fibroblasts. Independently, physical contact and paracrine signals from macrophages differentially activated VF fibroblasts.27 To that end, understanding interactions between VF fibroblasts and macrophages is foundational to optimally treat VF disease.
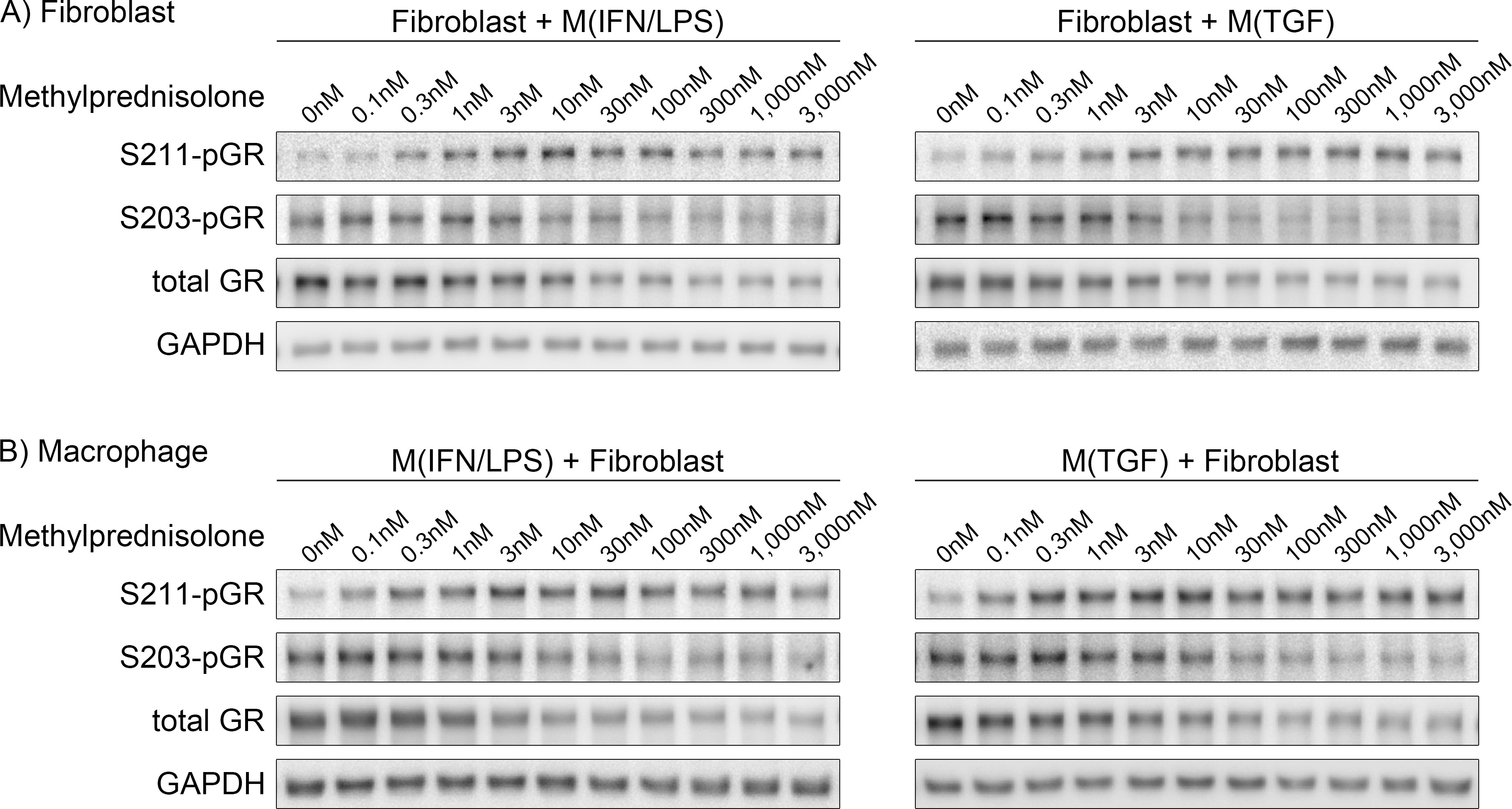
Recently, refinement of GC dose has emerged as a possible strategy to improve GC therapy.15,30 Our previous work with indirect co-culture models found fibrotic and inflammatory responses of VF fibroblasts triggered by macrophage-derived paracrine signals were promoted and inhibited by ‘high’ and ‘low’ concentrations of methylprednisolone, respectively.31 Based on this finding, we hypothesized minimizing GC concentrations to sufficiently inhibit inflammation improves efficacy of GC therapy. However, previous co-culture studies employed a cell culture insert to allow only paracrine signaling. Considering the in vivo environment in which macrophages directly engage fibroblasts,32 co-culture models with direct intercellular communication further support the translation of in vitro findings to support in vivo investigation. In addition, mechanisms underlying concentration-dependent negative and positive gene regulation remain unknown.
Based on the currently known biochemistry of GC signaling, unrelated to concentration-dependency, complex reactions of the GC receptor (GR) are thought to be a source of diverse GC functions.10,33 GR interacts with numerous proteins. The GC/GR complex binds and inhibits other transcription factors in the cytoplasm. Alternatively, GR translocated to the nucleus binds to both negative and positive gene regulatory elements. Various post-translational modifications (phosphorylation, acetylation, SUMOylation) are involved in GR distribution and recruitment to gene regulatory elements. Additionally, accessibility to negative and positive gene regulatory elements is putatively altered by dimerization of GR concentrated in the nucleus.10
Despite diversity of GR interactions with other signaling pathways,11 recent transcriptomic analysis on VF fibroblasts highlighted the impact of GR on the Hippos signaling pathway, which has a key role in fibrosis.34–37 In this pathway, Yes-associated protein (YAP) and transcriptional co-activator with PDZ-binding motif (TAZ) are the core.38 Activated YAP/TAZ enters the nucleus and primarily serves as co-activators of TEA domain transcription factors (TEADs) to induce TEAD-dependent transcription. CCN2, a fibrotic gene induced by high-concentration GCs, is a target of YAP/TAZ-TEAD signaling;39 this finding underlies the hypothesis that YAP/TAZ-TEAD signaling is specifically activated by high-concentration GCs.
In the current study, a direct co-culture model was employed to further confirm concentration-dependent effects of methylprednisolone to alter fibrotic and inflammatory responses of human macrophages and VF fibroblasts. We additionally explored nuclear localization of GR and YAP/TAZ in this model to interrogate mechanisms underlying concentration-dependent effects of GCs. Ultimately, we seek to refine GC therapy corroborated by mechanistic insight, to benefit millions of patients with voice-related disability.
{kind=link}