4.1 Comparative anatomy of the intrinsic scapular and shoulder muscles in procyonids
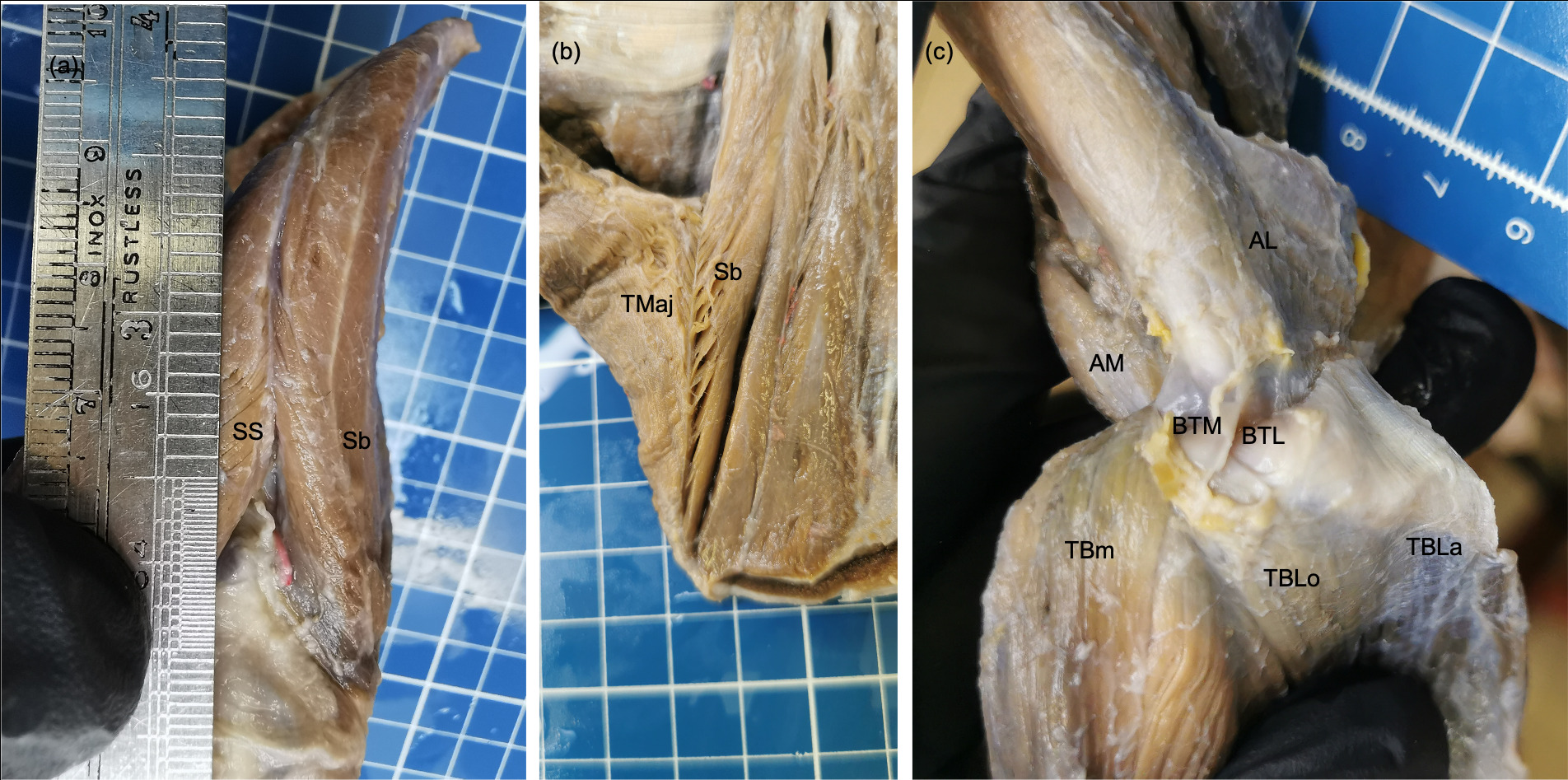
The m. deltoideus of P. cancrivorus had a similar arrangement to that described formerly in the same species (Santos et al. 2010b; Tarquini et al. 2023), and other procyonids, such as Procyon lotor (Allen 1882; Feeney 1999), Nasua nasua (Mackintosh 1875; Santos et al. 2010a; Böhmer et al. 2020; Tarquini et al. 2023), Nasua narica (Mackintosh 1875), Bassaricyon alleni (Beddard 1900), and P. flavus (Beswick-Perrin 1871; Windle and Parsons 1897; Böhmer et al. 2020). However, the pars scapularis always originated from the fascia of the m. infraspinatus, in contrast with the findings of Tarquini et al. (2023) who only found that origin in N. nasua. On the other side, the variant origin of the pars scapularis from a common aponeurosis with the m. teres minor was not reported in any procyonid.
The extended origin cranially to the cranial margin of scapula of the supraspinatus and subscapularis muscles forming an intermuscular septum had been described similarly in the same species (Santos et al. 2010b)and N. nasua (Santos et al. 2010a). The origin between both muscles was not reported in other studies in the same species (Windle 1888; Tarquini et al. 2023), and neither in P. lotor (Allen 1882; Windle and Parsons 1897)and P. flavus (Beswick-Perrin 1871; Windle and Parsons 1897; Böhmer et al. 2020; Vélez-García et al. 2023). In one specimen of N. nasua, a strong fascia was between both muscles, and in another specimen both muscles shared fibers (Tarquini et al. 2023). The variant presence of two bellies in the m. supraspinatus also was reported in this species (Tarquini et al. 2023), but in that case, the accessory belly was superficial but not located cranially, such as occurred in one P. cancrivorus specimen of the present study and one P. lotor(Allen 1882). The insertion onto the humeral transverse retinaculum was found in none, while Tarquini et al. (2023) found it in their three specimens.
The m. teres minor was completely independent to the m. infraspinatus in P. cancrivorus, such as was formerly described in the same species (Windle 1888; Pereira et al. 2010; Santos et al. 2010b; Tarquini et al. 2023) and other procyonids (Davis 1949; Böhmer et al. 2020; Tarquini et al. 2023; Vélez-García et al. 2023). In some specimensof P. lotor and P. flavus, the muscle may be fused to the m. infraspinatus (Beswick-Perrin 1871; Allen 1882; Julitz 1909). The origin of the m. teres minor from the infraglenoid tubercle as was reported in P. cancrivorus and N. nasua (Santos et al. 2010a, b) was not found since the tubercle was occupied by the origin tendon of the m. triceps brachii caput longum, such as was recently reported (Tarquini et al. 2023). The origin of the m. infraspinatus from the origin aponeurosis of the m. teres minor has only been described in P. flavus (Vélez-García et al. 2023).
Based on the most authors, the m. subscapularis does not have important differences among procyonid species (Beswick-Perrin 1871; Mackintosh 1875; Windle and Parsons 1897; Julitz 1909; Santos et al. 2010a, b; Böhmer et al. 2020). However, it is divided into two portions in Bassariscus (Davis 1949), P. lotor (Davis 1949), N. narica (Davis 1949), and N. nasua (Tarquini et al. 2023). Besides, the muscle does not only originate from the subscapular fossa but from the cranial and caudal margins of the scapula in P. cancrivorus (Tarquini et al., 2023; present study)and N. nasua (Tarquini et al. 2023).
The origin of the m. teres major from the infraspinatus and subscapularis muscles contrasts with other studies that did not find a connection with these muscles in the same species (Santos et al. 2010b; Tarquini et al. 2023). While in P. lotor (Allen 1882; Davis 1949), B. astutus, N. narica (Davis 1949) and N. nasua (Tarquini et al. 2023), the origin is very similar to that found in our P. cancrivorus specimens. The origin from the infraspinatus muscle is not found in P. flavus (Windle and Parsons 1897; Julitz 1909; Vélez-García et al. 2023), and in some specimens of N. nasua (Santos et al. 2010a). Other authors did not report none origin to this muscle in P. cancrivorus (Windle 1888) and Nasua (Mackintosh 1875). On the other side, the insertion onto the humerus was not found separated from the m. latissimus dorsi as was reported by other authors (Tarquini et al. 2023).
The m. coracobrachialis of P. cancrivorus presented a small shape, such as was reported formerly in the same species (Windle 1888; Tarquini et al. 2023), and other procyonids (Mackintosh 1875; Allen 1882; Beddard 1900; Santos et al. 2010a; Tarquini et al. 2023). P. flavus has another m. coracobrachialis named m. coracobrachialis longus (Beswick-Perrin 1871; Windle and Parsons 1897; Julitz 1909; Vélez-García et al. 2023), which is absent in P. cancrivorus and other procyonids. However, it can also be absent in some P. flavus specimens (Vélez-García et al. 2023).
4.2 Comparative anatomy of the brachial muscles in procyonids
The m. biceps brachii had one head in most specimens of P. cancrivorus, being similar to that formerly described (Mackintosh 1875; Windle 1888; Pereira et al. 2010; Santos et al. 2010a, b; Tarquini et al. 2023). The other insertion onto the ulnar tuberosity described by some authors (Pereira et al. 2010; Santos et al. 2010a, b) was not found by us and other studies (Windle 1888; Tarquini et al. 2023). In P. flavus, the presence of a well-developed second head (caput breve) originating from the coracoid process of scapula is normal (Beswick-Perrin 1871; Windle and Parsons 1897; Julitz 1909; Böhmer et al. 2020; Vélez-García et al. 2023). In P. lotor, that same head may be present in a feeble shape as anatomical variant (Windle and Parsons 1897). However, that head (caput breve) is not the same accessory head found in one P. cancrivorus specimen, since it originated from the humerus. Similarly, a small humeral head was found in one case of P. flavus (Vélez-García et al. 2023).
The m. brachialis originated from along caudolateral surface of the humerus including the medial aspect of the lateral supracondylar crest in P. cancrivorus, which is similar to that reported previously (Tarquini et al. 2023). In contrast, other studies reported the origin only from the proximal part of the humerus in the same species (Pereira et al. 2010; Santos et al. 2010b) and N. nasua (Santos et al. 2010a; Böhmer et al. 2020). The origin from the whole lateral surface of the humerus was described in N. nasua (Mackintosh 1875), N. narica (Mackintosh 1875), P. lotor (Allen 1882), and P. flavus (Julitz 1909; Vélez-García et al. 2023). However, the origin has been described from the proximal half of the humerus in the latter species (Beswick-Perrin 1871). The insertion only onto the ulna agrees with that described by most authors in procyonids (Beswick-Perrin 1871; Mackintosh 1875; Allen 1882; Windle 1888; Beddard 1900; Julitz 1909; Böhmer et al. 2020; Tarquini et al. 2023), while the insertion onto the radius described by some authors in P. cancrivorus and N. nasua was not found (Pereira et al. 2010; Santos et al. 2010a, b).
The four heads of the m. triceps brachii in P. cancrivorus were found as was previously reported by other authors (Pereira et al. 2010; Santos et al. 2010b), and similar to B. alleni (Beddard 1900), P. lotor (Allen 1882; Feeney 1999), N. nasua (Santos et al. 2010a), and P. flavus (Julitz 1909; Vélez-García et al. 2023). Several studies did not describe the caput accessorium of the m. triceps brachii or a homologous portion in procyonids (Beswick-Perrin 1871; Mackintosh 1875; Windle 1888; Böhmer et al. 2020). Former studies only reported three heads in P. cancrivorus (Windle 1888), N. nasua (Mackintosh 1875), and P. flavus (Beswick-Perrin 1871). A more recent study named four heads in N. nasua and P. flavus (Böhmer et al. 2020), although the caput mediale accessorium is actually the m. anconeus medialis (Vélez-García et al. 2023). In another more recent study was described that the muscle has five heads in P. cancrivorus and N. nasua (Tarquini et al. 2023), although, the caput mediale accessorium is actually the m. anconeus medialis. P. lotor and P. flavus may have five heads due to the presence of a second caput laterale (Windle and Parsons 1897; Vélez-García et al. 2023). N. narica may have four heads due to the presence of a second caput longum from the edge of the glenoid cavity (Mackintosh 1875), which could be similar to that found in P. cancrivorus where the caput longum had two origins. In our study, several differences were found concerning that reported by Tarquini et al. (2023). Among them, fleshy fibers were not found at the origin of the caput laterale; the caput laterale sent fleshy fibers to all other heads; two tricipital bursas; and the division of the caput longum tendon by a perforating branch of the subscapular artery.
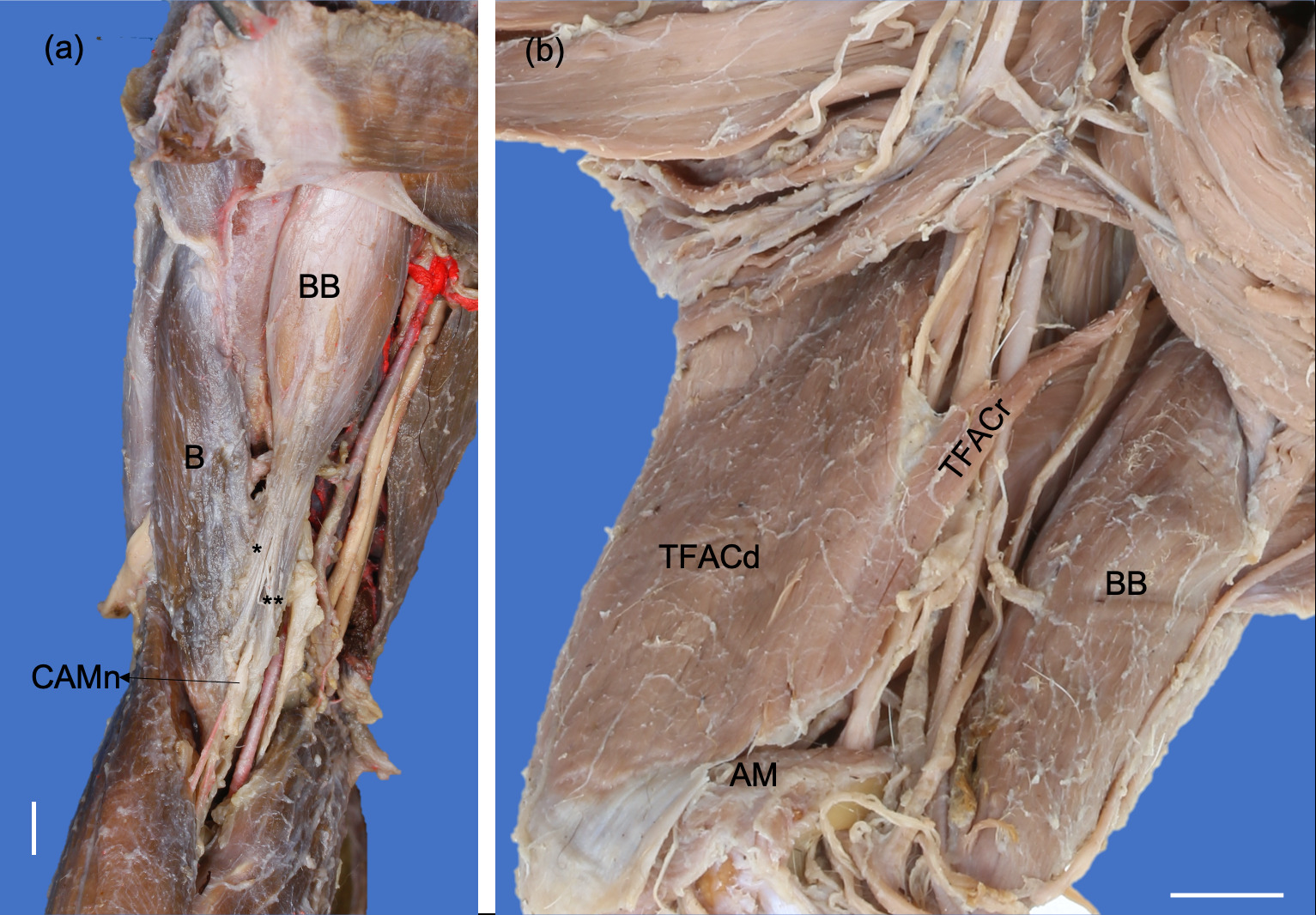
The formation of two parts (cranial and caudal parts) in the m. tensor fasciae antebrachii in P. cancrivorus has only been described in P. lotor (Feeney 1999), P. flavus (Vélez-García et al. 2023) and B. alleni (Beddard 1900). The origin from the m. teres major was only found in N. narica (Mackintosh 1875) and P. flavus (Vélez-García et al. 2023). The insertion onto the olecranon and antebrachial fascia was reported in P. flavus (Beswick-Perrin 1871; Julitz 1909; Vélez-García et al. 2023) and one study in P. cancrivorus (Pereira et al. 2010). While the unique insertion onto the olecranon was described in most procyonids (Mackintosh 1875; Allen 1882; Santos et al. 2010a; Böhmer et al. 2020; Tarquini et al. 2023). The tendinous fusion with the caput longum tendon of m. triceps brachii was not reported in any other study.
The m. anconeus medialis in P. cancrivorus had a similar arrangement to that described in a homologous part (with another term or portion of the m. triceps brachii) in the same species ("caput mediale accessorium" Tarquini et al., 2023), N. nasua ("m. triceps brachii caput mediale" Böhmer et al., 2020; Mackintosh, 1875), P. lotor ("m. anconeus epitrochlearis" Allen, 1882; Windle & Parsons, 1897), and P. flavus (Beswick-Perrin 1871; Windle and Parsons 1897; Vélez-García et al. 2023). We found origin also from the medial epicondyle of the humerus in all P. cancrivorus specimens similar to that found in P. flavus (Vélez-García et al. 2023). In contrast, it differs from that reported in the same species and N. nasua where the origin only was from the supracondylar foramen (Tarquini et al. 2023). This muscle or a homologous portion was not described by other authors (Windle 1888; Pereira et al. 2010; Santos et al. 2010b), however, in one of those studies, it was pointed as accessory head of the m. triceps brachii in the figure 3 (Santos et al. 2010b). This corroborates that the muscle several times is missed by the authors, which may be because it is not present at the NAV (ICVGAN 2017). In the case of N. narica, both anconei muscles are reported united to the biceps (Mackintosh 1875). However, this union could be a mistake of the author and could have referred to the m. triceps brachii since the anatomical relationship is closer with this muscle than the m. biceps brachii.
The origin of the m. anconeus lateralis (m. anconeus) extended proximally reaching part of the humeral shaft in our specimens of P. cancrivorus, which differed from other studies where the muscle only originates from the lateral supracondylar crest (Mackintosh 1875; Pereira et al. 2010; Santos et al. 2010b; Böhmer et al. 2020; Tarquini et al. 2023). P. lotor (Allen 1882) and P. flavus (Vélez-García et al. 2023) are the only two procyonid species where the muscle reaches the humeral shaft, even being more proximally extended than P. cancrivorus. The muscle was not reported in a former study of P. cancrivorus (Windle 1888). In some specimens of P. flavus, the m. anconeus lateralis may be fused to the caput mediale of the m. triceps brachii (Beswick-Perrin 1871; Julitz 1909).
4.3 Anatomical variants of P. cancrivorus present in other carnivorans
Some anatomical variants found in P. cancrivorus may be present in other species within the order Carnivora. The presence of two bellies in the m. supraspinatus has been reported in the canid Cerdocyon thous (Vélez-García et al. 2018b), the felid Panthera leo (Barone 1963), and the viverrid Civettictis civetta (Macalister 1873b). While in the mustelid Galictis cuja, it has three bellies (Ercoli et al. 2015). The origin of the m. infraspinatus from the aponeurosis of the m. teres minor has been described in the ailurid Ailurus fulgens (Fisher et al. 2009).
The m. tensor fasciae antebrachii has only been reported with more than one part in a few species. In two species, it has been reported with two portions with similar origins to those of P. cancrivorus, such as the mustelids Eira barbara (Macalister 1873), Martes caurina, Martes martes (Yousefi et al. 2018), Pekania penanti (Feeney 1999), and the ursid Ailuropoda melanoleuca (Davis 1964). In the ursid Ursus americanus, it was reported with three portions and inserted onto the medial epicondyle and olecranon (Shepherd 1883). The insertion onto the triceps brachii tendon by the m. tensor fasciae antebrachii has been reported in the canid Canis lupus familiaris (Hermanson 2020), the mustelid M. martes (Yousefi et al. 2018) and the felid Leopardus pardalis (Julik et al. 2012). The origin from the m. teres major by the m. tensor fascia antebrachii has been reported in U. americanus (Shepherd 1883), M. martes (Yousefi et al. 2018) and the felid Puma concolor (Concha et al. 2004).
From our knowledge, two anatomical variants found in P. cancrivorus have not been reported in other carnivorans, such as the common origin aponeurosis of the teres minor and deltoideus pars scapularis muscles; the two variations of the m. biceps brachii; and the fusion of the laterale and accessorium heads of the m. triceps brachii.
4.4 Comparative functional and evolutionary analysis of the intrinsic shoulder and brachial muscles in Procyon cancrivorus based on the topology and innervation
The functional analysis has been excellently developed in the study of Tarquini et al. (2023) since they compared the muscle volume among muscular groups and other species. However, below, we include other functional and evolutionary inferences that were not analyzed in P. cancrivorus.
The m. deltoideus in procyonidsis mainly divided into two parts, however, based on the topology of the m. cleidobrachialis and the distribution of the axillary nerve (Enciso-García and Vélez-García 2022; Vélez García et al. 2023), this latter muscle is the third part of the m. deltoideus named pars clavicularis (Vélez-García and Miglino 2023). Evolutionarily, it was a part joined to the pars acromialis from the reptiles to the last common ancestor of mammals ("deltoideus acromialis et clavicularis" Diogo et al., 2016). In carnivorans, both parts should have been separated due to the involution of the clavicle and its functional antagonism. This is because the acromialis and scapularis parts act together to flex the shoulder, and the pars clavicularis act to extend the shoulder together the m. cleidocephalicus (Diogo et al. 2012; Hermanson 2020; Vélez-García and Miglino 2023; Vélez‐García et al. 2023). The origin in common with the m. deltoideus pars scapularis and m. teres minor in one limb of P. cancrivorus could be associated with the evolutionary derivation of the m. teres minor from the m. deltoideus in tetrapods (Diogo et al. 2018, 2019). This can be supported within carnivorans since the m. teres minor has been found fused to the m. infraspinatus in mustelids of the superfamily Lutrinae (Otters) (Macalister 1870; Howard 1973; Ramírez Arango et al. 2024). Recently, it was corroborated due to the caudal aspect of the m. infraspinatus was being innervated by the n. axillaris in Lontra longicaudis (Ramírez Arango et al. 2024). Therefore, the presentation of a common origin of the deltoideus pars scapularis and teres minor muscles is a phylogenetic trade associated with the muscular derivation of the shoulder flexor muscles in mammals (Table 2).
The shoulder joint is extended and stabilized by the supraspinatus and infraspinatus muscles. Besides, the presence of an intermuscular septum between the supraspinatus and subscapularis muscles in carnivorans has been related to a more force to extend and stabilize the shoulder since the bands of both muscles are disposed cranially to the scapula. This arrangement has been associated in species with locomotion mainly cursorial as canids (Vélez-García et al. 2018b) and as occurs in P. cancrivorus. The m. biceps brachii is another muscle that supports cranially the shoulder joint and extends it since its tendon of origin crosses internally the joint capsule of the shoulder.
The teres major, teres minor, and subscapularis are shoulder flexors, and they are powered by the caudal portion of the m. tensor fasciae antebrachii due to its caudal arrangement together with the caput longum of the m. triceps brachii in P. cancrivorus. The same muscle support should occur in other carnivorans where the m. tensor fasciae antebrachii has a caudal arrangement, such as other procyonids (Beswick-Perrin 1871; Julitz 1909; Böhmer et al. 2020; Vélez-García et al. 2023), most mustelids (Macalister 1873; Cohen and Hart 1968; Howard 1973; Leach 1977; Ercoli et al. 2015; Böhmer et al. 2018, 2020; Yousefi et al. 2018; Ramírez Arango et al. 2024), ursids (Shepherd 1883; Davis 1964), and some felids (Concha et al. 2004; Vargas et al. 2017).
The m. coracobrachialis is adductor and extensor of the shoulder(Hermanson et al. 2020). The presence of only one m. coracobrachialis could be associated with species with more cursorial abilities that do not need major adduction force (Vélez-García et al. 2023), such as occurs in P. cancrivorus. The presence of two coracobrachialis muscles is a feature only present in some species of the suborders Caniformia and Feliformia. Among caniforms, this character is present in the procyonid P. flavus (Beswick-Perrin 1871; Julitz 1909; Vélez-García et al. 2023), mustelids of the genera Martes, Pekania and Eira (Macalister 1873; Mackintosh 1875; Leach 1977; Yousefi et al. 2018), the ailurid Ailurus fulgens (Carlsson 1925; Fisher et al. 2009), and ursids (Shepherd 1883; Kelley 1888; Windle and Parsons 1897; Davis 1964; Annie et al. 2019). Among feliforms, it is present in the euplerid Cryptoprocta ferox (Carlsson 1925; Böhmer et al. 2020)and viverrids of the genus Genetta (Taylor 1982)(Figure 7). The presence of two coracobrachialis muscles is phylogenetically related from the amphibians (Diogo et al. 2018), which is an arrangement that persists in reptiles, monotremes, some marsupials, and some eutherians (Gambaryan et al. 2015; Diogo et al. 2018; Richards et al. 2023). However, from an analysis of the functional and phylogenetic point of view within the family Carnivora, most of these species with two coracobrachialis muscles could be associated with a common ancestor with high arboreal abilities (Figure 7). This is supported due to the m. coracobrachialis has been lost together with m. teres minor in mustelids that do not have any arboreal ability, such as occurs in otters (Lutrinae) who are more specialized for natatorial locomotion (Haughton 1864; Windle and Parsons 1897; Cohen and Hart 1968; Howard 1973; Ramírez Arango et al. 2024).
The elbow flexion is performed mainly by the biceps brachii and brachialis muscles, besides during flexion, they are supported by the brachioradialis and extensor carpi radialis muscles (Hermanson 2020; Vélez-García et al. 2022). Therefore, the presence of a second head in the m. biceps brachii should potentiate the elbow flexion intraspecifically in P. cancrivorus due to an origin more distal since it originated from the humerus. In other carnivorans with caput breve, the elbow flexion should increase the flexion velocity of the elbow since it has a more proximal origin (scapula), such as occurs in P. flavus (Böhmer et al. 2020; Vélez-García et al. 2023), A. fulgens (Fisher et al. 2009), and some ursids (Shepherd 1883; Kelley 1888; Davis 1964). The presence of a vestigial caput breve of m. biceps brachii in a P. lotor (Windle and Parsons 1897) could support the hypothesis that the common ancestor of procyonids potentially had it and conserved it from the common ancestor of arctoids. The caput breve could have disappeared within the family Procyonidae when the non-Potos genera diverged from the genus Potos (Table 2) (Fig. 7). Previously, the presence of several bicipital arteries had been associated with a higher activity of the m. biceps brachii in P. flavus (Vélez-García et al. 2023). However, after reviewing the arterial supply to the m. biceps brachii in P. cancrivorus, the presence of several bicipital arteries could be a characteristic within procyonids, which differs from canids (Vélez et al. 2018; Hermanson et al. 2020) and Felis catus (ICVGAN 2017), which only have one bicipital artery.
The elbow extension in P. cancrivorus is powered by a m. triceps brachii with four heads, two anconeal muscles, and the two parts of the m. tensor fasciae antebrachii. Tarquini et al. (2023) for P. cancrivorus reported the cranial part of the m. tensor fascia antebrachii as a “caudal belly” to this, which originated from the m. cutaneous trunci and was also considered as part of this latter. However, these authors did not take into account the innervation and the direction of the muscle fibers to this belly, which should be reviewed to infer muscle derivation in vertebrates (Diogo and Abdala 2010). Therefore, based on the radial nerve distribution where the branch to the caudal part branched also to the cranial part, and the direction of the fibers is parallel, said belly is actually a part of the m. tensor fasciae antebrachii (Figure 4). Furthermore, the insertion aponeurosis is also fused to the tendon of the m. triceps brachii caput longum (Figure 6b), which is not reported in other studies of the same species and other procyonids. Thus, the synapomorphy of several origins in arctoids for the m. tensor fasciae antebrachii is also retained in P. cancrivorus. Previous studies in caniforms have determined that canids only conserve the cranial part of the m. tensor fasciae antebrachii while arctoids the caudal part (Feeney 1999; Vélez-García et al. 2023). In contrast, based on the origin of the m. tensor fasciae antebrachii only from the m. latissimus dorsi in canids (Feeney 1999; Pereira et al. 2016; Souza-Junior et al. 2018; Vélez et al. 2018; Böhmer et al. 2020; Smith et al. 2020), actually the part more conserved in caniforms is the caudal part while the presentation of the cranial part is variable in arctoids. What happens is that the m. tensor fasciae antebrachii is so developed in arctoids that it extends caudally to the caput longum of the triceps brachii, and is observed in the lateral views of the figures of several studies (Hall 1926; Davis 1964; Fisher et al. 2009; Moore et al. 2013; Ercoli et al. 2015; Böhmer et al. 2020; Tarquini et al. 2023; Vélez-García et al. 2023; Ramírez Arango et al. 2024). Besides, based on its topology and innervation, we could infer that this muscle is derived from the caput longum of the m. triceps brachii in carnivorans, which agrees with the evolutionary derivation from the last common ancestor of mammals (Diogo et al. 2018). The presence of a caput accessorium in the m. triceps brachii has been found in monotremes, and it has been inferred that it is a division of the caput mediale caused by the pass of the radial nerve (Gambaryan et al. 2015), which also occurs in carnivorans. On the other hand, the fusion presented by the laterale and accessorium heads in one P. cancrivorus specimen, and the distribution of the radial nerve to these heads, allow us to suggest that the caput accessorium could also be evolutionarily derived from the caput laterale in procyonids (Table 2).
The extensor elbow muscles are supplied by the caudal circumflex, deep brachial, radial, and ulnar collateral arteries in P. cancrivorus, similar to that described in P. flavus (Vélez-García et al. 2023), A. melanoleuca (Davis 1964), C. thous (Vélez et al. 2018) and C. lupus familiaris (Hermanson et al. 2020). In PcS6, the deep brachial artery originated from the thoracodorsal artery differing from other specimens., Similarly in A. melanoleuca, that branch was described as a descending branch of the thoracodorsal artery that anastomoses with the deep brachial artery (Davis 1964). This differs from that presented in one specimen of P. cancrivorus since there was no formation of a homologous branch from the brachial artery. Unilaterally in PcS1, the deep brachial artery was formed by two branches of the brachial artery, which is not reported in any species.
Based on the former dissections performed by Windle and Parsons (1897) in several carnivorans, the m. anconeus medialis is the most constant and is supplied by the ulnar nerve. This has been corroborated more recently in procyonids (Enciso-García and Vélez-García 2022; Vélez García et al. 2023), the mustelid Lontra longicaudis (Ramírez Arango et al. 2024), and felids (Barone 2020a; Barreto‐Mejía et al. 2022). In contrast, this muscle normally is not present in canids (Pereira et al. 2016; Souza-Junior et al. 2018; Vélez et al. 2018; Hermanson 2020; Smith et al. 2020) and ursids (Shepherd 1883; Davis 1964). If it is present in those species, it has a vestigial shape (Kelley 1888; Vélez-García et al. 2018a; Böhmer et al. 2020) or is fused to the caput mediale of the m. triceps brachii as was reported in the ursid Ailuropoda melanoleuca (Davis 1964). Therefore, the m. anconeus medialis lost functionality in the families Canidae and Ursidae. Several myological studies have named this muscle as another head of the m. triceps brachii in species of the order Carnivora, such as those studies performed in caniforms (Shepherd 1883; Leach 1977; Fisher et al. 2009; Ercoli et al. 2015; Böhmer et al. 2020) and feliforms (Julik et al. 2012; Viranta et al. 2016; Böhmer et al. 2020; Smith et al. 2021; Dunn et al. 2022). However, due to its topology and innervation by the ulnar nerve, the m. anconeus medialis in the order Carnivora retains its evolutionary derivation together the m. flexor carpi ulnaris in the caudolateral muscular complex of the antebrachium (Table 2) (Figure 7). This is due to the muscle is present in chordates from the amphibians, and it is not derivate from the m. triceps brachii (Diogo and Abdala 2010; Diogo et al. 2018; Molnar and Diogo 2021). The m. anconeus medialis should also act as a medial stabilizer of the elbow joint when the thoracic limb is fixed on the substrate. The presence of this muscle in procyonids could be related to a more developed medial epicondyle of the humerus (Tarquini et al. 2019; Vélez-García et al. 2023), which also gives origin to the caudomedial antebrachial musculature (McClearn 1985; Perdomo-Cárdenas et al. 2021). Therefore, high handling abilities in P. cancrivorus may also require more medial stabilization of the elbow while the caudomedial antebrachial musculature is acting.
In conclusion, the most intrinsic shoulder and brachial musclesof P. cancrivorus potentially conserve the evolutionary derivation of the last common ancestor of mammals based on the topology, innervation, and anatomical variants. However, the division of the m. tensor fasciae antebrachii into two parts is a characteristic that appears within the infraorder Arctoidea and remains in most cases in P. cancrivorus.
{kind=link}
{kind=link}