In most populations, 3–5% of PD is explained by genetic causes linked to known PD genes. However, the majority of patients (90–95%) have idiopathic or sporadic PD [26]. Further, epidemiological studies indicate a relationship between various environmental factors such as pesticide exposure and the development of sporadic PD in later stages of life [1, 27]. Hence, we developed an adult life stage (health and transition phase) specific fly model of PD using mitochondrial complex I inhibitor neurotoxicant rotenone [5]. Adult life phase specific gene expression profile variation across model organisms suggest the relevance and importance of life phase-specific animal models for understanding the pathophysiology of late-onset NDDs such as PD and screening small molecules/nutraceuticals with potential neuroprotective efficacy and deciphering molecular basis of their biological activity.
Studies on animal models have shown the neuroprotective efficacy of CUR [16–18]. Contradictory reports of concentration-dependent CUR toxicity have been reported in animal models [28] and in Drosophila model [23]. However, the toxicity of CUR in animal models is under-reported. Hence, we performed a comprehensive experiment with an array of CUR concentrations to identify the non-toxic concentrations of CUR in both the adult early health phase and transition phase of the Drosophila.
Present experimental approach employing CUR was to expose the Drosophila to a range of concentrations to determine its detrimental effects and select a suitable range of non-toxic concentrations for further studies. We employed 8 concentrations ranging from 25 µM to 2.5 mM of CUR in both the adult early health phase and transition phase male Drosophila. It was found that CUR concentrations of 2.5 mM had a deleterious effect on the viability of the fly, whereas concentrations lower than 2.5 mM showed no observable toxicity in the early adult health phase fly up to 10 days (Fig. 2). In the highest concentration of 2.5 mM, all the flies employed in the experiment were dead by the 30th day and 11th day of CUR treatment in the case of the early health and transition phases respectively. The concentration below 1 mM did not show any mortality up to 10 days of exposure to CUR, indicating that there was no toxicity in all the lower concentrations. CUR toxicity study in human also suggests that it primarily causes gastrointestinal disturbances, diarrhoea, as well as distension and gastroesophageal reflux disease [29]. Therefore, it is essential to make out the toxic concentrations of CUR before employing it as a neuroprotective agent in the fly model of PD. Hence, we picked up sub-lethal concentrations of CUR (100 µM, 250 µM, 500 µM and 1 mM) and the same concentrations were employed in life phase-specific ROT-mediated Drosophila model of PD under pre- and co-feeding regimen.
To understand DA-ergic neurodegeneration and to decipher the effectiveness of therapeutic molecules, many laboratories either co-treat or pre-treat young PD fly models belonging to the health phase to screen small molecules/drugs/nutraceuticals and concluded about their DA-ergic neuroprotective efficiency [6, 21, 22, 30]. They determined the neuroprotective efficacy of the molecules by assessing behavioral markers such as mobility defects, biochemical markers such as anti-oxidant enzyme levels and levels of brain DA and its metabolite levels, or cytological markers such as degeneration of DNs in the whole brain of young animals whereas late-onset NDDs such as PD onsets during the transition phase of adult life!!
The present study indicates that feeding the fly of the adult early health phase with ROT alone adversely decreases mobility (~ 30%) which could be improved upon co-feeding with CUR. Feeding the fly with CUR per se did not show any mobility defects (Fig. 3A & B). Results indicate that in both the co-feeding (Fig. 3A & B) and pre-feeding regimens (Fig. 3C-H), CUR could confer neuroprotection during the early health phase. It has been reported that exposure to ROT induces severe mobility defects in Drosophila and the same can be rescued upon co-feeding with CUR [30]. In the case of the late health phase fly, CUR fails to rescue the mobility defects in both the co- and pre-feeding regimens (Suppl. Fig. S2). In the case of the transition phase fly, CUR fails to rescue ROT-mediated mobility defects in both the co- and pre-feeding regimens as determined through the negative geotaxis assay (Fig. 4). These results are in agreement with previous work from our lab, where CUR confers DA-ergic neuroprotection only during the early health phase but not during the transition stage [23]. However, in the present study, CUR fails even during the late health phase, which can be attributed to mitochondrial complex I inhibitor ROT specific neurotoxicity. Previous studies demonstrated the neuroprotective efficacy of CUR in ROT-mediated neurotoxicity in cell culture, young adult Drosophila and rat models and substantiated that CUR sequesters the intracellular and mitochondrial ROS levels and inhibits the caspase-3/caspase-9 activity [16, 22, 31, 32]. These results suggest that CUR may be a potential therapeutic compound for PD intervention. However, all these studies used young animals belonging to the adult early health stage. The results from the present study suggest that CUR can rescue ROT-mediated mobility defects in ALSS fashion; meaning CUR confers neuroprotection only during the early health phase and not in the late health phase and transition phase of the adult life.
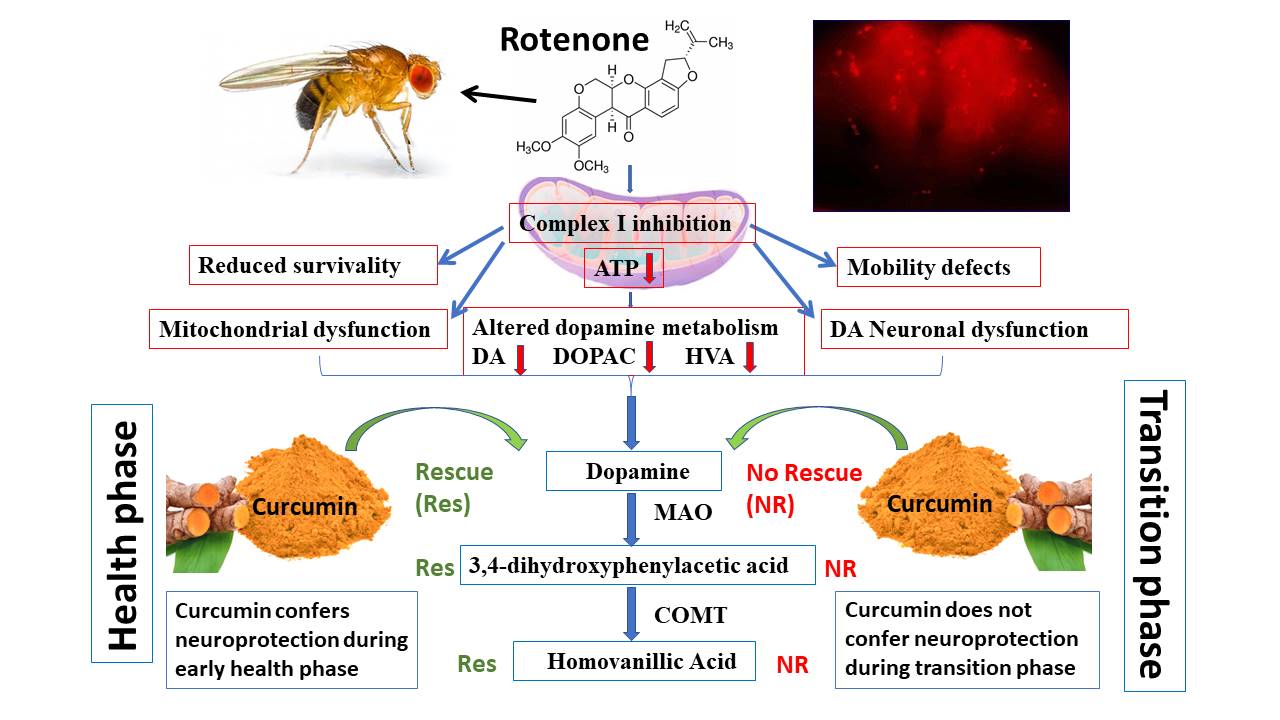
The death of DNs is the characteristic pathological marker of PD. Hence, in order to decipher the extent of DA-ergic neurodegeneration under induced PD conditions and possible rescue by CUR, we quantified the DNs in the whole fly brain followed by characterization of DA “neuronal dysfunction” by quantifying the FI emanated from the fluorescently labelled secondary antibody targeted against the primary anti-TH antibody using fluorescence microscopy. The results illustrate that there is no variation in the number of DNs between the control and PD brains of Drosophila in both the health and the transition phases of adult life (Fig. 5B & 6B). This observation is consistent with the previous works from our lab [5] and other laboratories [25, 33, 34], which is logical in the light of “dying back” phenomenon. The issue of loss of DNs per se in fly brain has been thoroughly evaluated in multiple fly models of PD (both genetic and sporadic) and demonstrated that there is no structural loss of DA neurons, but only the synthesis of TH is diminished [25]. Hence, we went ahead with the quantification of the FI of TH-positive neurons. Results illustrate that the fly treated with ROT alone led to a significant decrease in FI to ~ 50% which could be significantly rescued by 25–30% upon co-feeding with CUR during the early health phase (Fig. 5C) while the diminished FI could not be rescued by CUR during the transition phase (Fig. 6C). We also analyzed the results by quantifying the FI of all the DNs present in the fly brain of different treatment groups (Fig. 5D & 6D). Similar results were obtained when analyzed in treatment group-wise. As the FI of the secondary antibodies is directly proportional to the level of synthesis of the TH, diminished levels of FI suggest the decreased levels of TH. Hence, it is possible that the level of DA synthesis might be reduced in the fly brain. This approach allowed us to accurately assess the extent of neurodegeneration of DNs in the fly PD models. This result is further substantiated by quantifying the levels of the brain-specific neurotransmitter DA and its metabolites (DOPAC and HVA) using HPLC-ECD.
The present study shows that feeding the fly with ROT alone led to a significant reduction in the levels of DA, DOPAC and HVA during the early health and the transition phases of Drosophila (Fig. 7). Similar results showing significant reduction in DA, DOPAC and HVA levels are indicated in 10 days old young fly [22], young mice [35], young wistar rat [36]. All these results indicate that ROT significantly downregulates the DA level under induced PD conditions.
The present study demonstrates that diminished levels of DA, DOPAC and HVA upon ROT treatment could be significantly rescued through co-feeding with CUR during the early health phase but not during the transition phase of the adult fly (Fig. 7). Study on young 12 weeks old albino wistar rat showed a significant reduction in DA and DOPAC levels upon ROT-mediated conditions which could be significantly improved upon co-feeding with CUR [36]. Another study indicated that feeding Danshensu (a plant extract; 60mk/kg for 28 days) could significantly improve the levels of DA, DOPAC and HVA in ROT-induced young 12 weeks old C57BL/6 mice indicating that the levels of DOPAC and HVA are proportional to DA content [35]. CUR is shown to restore the levels of DA, DOPAC and HVA in neurodegeneration mediated by lipopolysaccharide in young Sprague-Dawley rats [37]. All these results support the present study and indicate that CUR restores the levels of DA, DOPAC and HVA in ROT-mediated PD models. However, these studies are restricted to 5–20 days young fly [6, 22] and 10–12 weeks young mice and rat models [35, 36] whose age is comparable with the early health phase of adult life. There is no data available relating to the levels of DA and metabolites in PD models of the transition phase of adult life, during which PD sets in. This is the first detailed study to have shown the levels of DA and its metabolites in the transition phase of Drosophila model of ROT-mediated PD.
Present study reveals that CUR does not influence the DA turn over during both the health and the transition phases of adult life. Research on DA metabolism gives an insight into the biochemical changes in DA-ergic system associated with PD. However, the biological significance of “turnover ratio” which denotes the level of metabolites (DOPAC and HVA) with respect to DA is not discussed in many studies quantifying DA and metabolites. Pagan et al. [38] reported DA metabolites as “exploratory biomarkers” with reference to CSF levels of DOPAC and HVA. Although, compiling the CSF levels of HVA with xanthine improves its use as a biomarker; CSF levels of HVA poorly correlate with PD severity or progression [39]. Though DOPAC is the primary metabolite of DA [40], LeWitt [41] commented that representing the CSF level of DOPAC as a marker for the progression of PD is not valid though it shows severity. A significant amount of DOPAC and HVA, albeit lower DA levels, is seen in the human brain wherein DA-ergic innervation has not been implicated [42]. Furthermore, the levels of neurotransmitters in CSF partially implicate the metabolism indicating limited relevance to neurotransmission in the brain. Hence, quantitative analysis of brain-specific DA and its metabolites concentrations are unrelated to DA-ergic neurotransmission which could be due to the possible role of capillary walls and glial cells in the catabolism of DA that needs to be further investigated [42]. Hence, the notion “turnover” is not applicable with respect to the activity of DA-ergic system [42].
These findings suggest that CUR-mediated nourishment of DA metabolism is restricted to the adult health phase and not to the transition phase. This illustration indicates that genetic targets and molecular networks of genotropic drug CUR-mediated correction process may not be active/expressive at optimum levels during later phases of the adult life. However, there is no significant difference in the DA turn over pattern in both the health and transition phase of the fly but trend shows a marginal slow-down in the catabolism of DOPAC to HVA during the transition phase of the fly (Fig. 8). Thus the study underlines the limitation of CUR as a therapeutic agent in late-onset NDDs such as PD. Figure 9 illustrates a detailed schematic presentation of our proposed study model.
The present study provides insights into the underlying reasons for neuroprotective efficacy of CUR during the adult health phase of Drosophila. Further, it also provides insights into the probable cause for the inefficacy of CUR during the adult transition phase of Drosophila. It is important to figure out why CUR is failing to sustain the activity of the TH during the transition phase, understanding of which will have critical implications in developing therapeutic strategies for late-onset neurodegenerative diseases such as PD. Figuring out the underlying molecular networks in an ALSS fashion will be a path-breaking contribution in addressing the pathophysiology of PD.
This is the first report to decipher the fact that CUR has limitations as a therapeutic molecule in the ROT-mediated idiopathic Drosophila model of PD, and its efficacy is restricted only to the early health phase of adult life.
{kind=link}