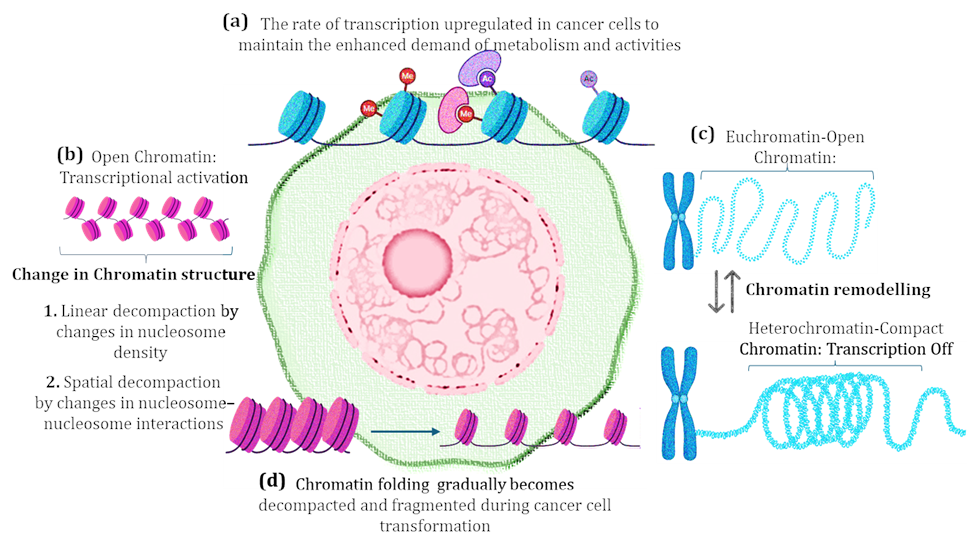
In this present study, super-resolution imaging of chromatin in normal vs cancer cell lines and quantitative analysis of the spatial distribution of domains across the nucleus of several breast cancer subtypes revealed that every cancer subtype has its different unique spatial distribution profile. Despite the importance of chromatin distribution and its positioning across the nucleus for proper gene expression, information about the chromatin spatial distribution profile in different diseases, and conditions especially in cancers remains extremely limited. The global distribution of chromatin across the nucleus can be shifted due to a reduction in heterochromatin density at a particular position (center to periphery and viceversa)28. It’s important to mention here that the formation of aberrant aggregates and the decrement of condensed dark nuclear material (heterochromatin) are the specific features of cancer cells8 Heterochromatin has an important role in chromosomal function and chromatin folding, due to which it participates in various human disease pathogenesis. Because heterochromatin can spread in support of transcription factors and has the potential to switch into facultative heterochromatin (works as euchromatin) in certain signals and is found to be frequently present in developmental genes29, 30. Based on these considerations we contemplate identifying the spatial distribution profile of chromatin, and euchromatin heterochromatin ratios in different breast cancer subtypes to target it as the prognostic marker and therapy. The spatial distribution profile (domain number, domain area, domain density, domain occupancy) of euchromatin revealed that the noncancerous cell line (HEK 293) exhibits a low number of nanodomains, domain area, domain density, and domain occupancy as compared to aggressive subtypes of breast cancer (Basal, Luminal-B, Luminal-H, and TNBC). This could be happened due to the increased rate of cell proliferation and gene expression in aggressive subtypes of breast cancer, as an elevated rate of gene expression requires more open chromatin structures 31. In contrast, the spatial distribution profile of euchromatin, and heterochromatin was found to be switched in several types of breast cancer variants. The noncancerous cell line (HEK 293) exhibits a high number of nanodomains, as compared to all the breast cancer subtypes except Luminal-H (SKBR-3 cell line). The reason behind this could be the exceptional types of chromatin remodeling in the case of Luminal-H subtypes, at this instance, the switching of H3K9me3 to H3K4me3 is taking place also the dispersion/decompaction of H3K9me3 is occurring to form several nanodomains. The lowest numbers of heterochromatin domains were observed in TNBC subtypes suggesting a high level of decompaction as compared to other breast cancer subtypes.
Further, the heterochromatin domain density was also found to be reduced in TNBC as compared to other breast cancer subtypes, and domain area and domain occupancy were also found to be reduced as compared to others. It should be mentioned that the Luminal-A subtype (MCF-7) was found to show a resemblance with TNBC in terms of domain area and domain occupancy. Further, we identified the euchromatin and heterochromatin ratios of all the breast cancer subtypes and compared them with non-cancerous HEK 293 cells and cervical cancer. Here also we identified that the ratio of euchromatin and heterochromatin has completely changed, and the amount of heterochromatin has drastically switched in TNBC. The H3K4me3: H3K9me3 ratio was also found to be switched in 4 out of 5 subtypes of the breast cancer subtypes but the highest switching rate was found in TNBC. Among them, the aggressive breast cancer subtypes displayed a high rate of switching. Based on this evidence we put forward that aggressive cancer subtypes (being hyperproliferative and hypertranscriptional) require an increased rate of gene expression which is supported by an increased rate of open chromatin structures and chromatin remodeling32. Likewise, this occurrence could be responsible for the decrement of heterochromatin (H3K9me3) in all the aggressive breast cancer cell lines8. The protein expression profiling of chromatin remodeling protein SSRP1 displays an elevated level of SSRP1 in all the cancer subtypes and supports that cancer cells' metabolic demand requires high gene expression and ultimately elevated chromatin remodelling (open chromatin). The high expression of H3K4me3 and H3K9me3 in Luminal A, Luminal H, and TNBC could be associated with the invasiveness of cancer, as upregulated H3K9me3 is previously reported in the invasive region of colorectal cancer33. Patients with breast cancer who have high levels of H3K4me3 in their tissue samples showed decreased survival rate33.
Based on the outcomes of our study we propose several factors that need to be investigated further (a) as we showed that euchromatin and heterochromatin ratios are changing in different breast cancers, what are those factors responsible for this, either histone chaperons, histone-modifying enzymes or some gene expressions are responsible? (b) Why H3K9Me3 domain’s density and occupancy were found to be diminished in MDA-MB-231 instead of high expression of H3K9me3 protein marks? (c) Either H3K9me3 (constitutive heterochromatin) converting to H3K27me3 (facultative heterochromatin) in aggressive breast cancer subtypes or heterochromatic decompaction is taking place?
Taken together our finding suggests the super-resolved imaging of chromatin (euchromatin and heterochromatin) and quantitative analysis of the spatial distribution of chromatin organization athwart the nucleus can distinguish the different higher-order organizations of chromatin in cancerous and non-cancerous cell lines. Our results underline that the spatial distribution of chromatin shifts after the cancer development, and the breast cancer subtypes Lumina A, B, H, basal, and triple-negative have distinct higher-order chromatin organization. The breast cancer subtype-specific cell lines MDA-MB-231(Triple negative) and SKBR-3 (Lumina-H) displayed the utmost euchromatin (H3K4me3) domain density, domain area, and domain occupancy and MDA-MB-231(Triple negative) showed nethermost heterochromatin (H3K9me3) domain density and occupancy instead of having overexpression of H3K9me3. The ratio of euchromatin and heterochromatin was also found to be distinct in MDA-MB-468, MCF-7, SKBR-3, and MDA-MB-231, as compared to reported euchromatin and heterochromatin packaging. Among all the non-cancerous and cancerous cell lines MDA-MB-231 displayed complete switching in euchromatin and heterochromatin ratios which suggests that MDA-MB-231 has an entirely distinct characteristic of euchromatin and heterochromatin spatial distribution among all the breast cancer subtypes which may be targeted to improve the breast cancer prognosis and risk stratifications. In addition, our finding also reveals elevated expression of SSRP-1 in all the cancer cell lines and overexpression of H3K4me3 and H3K9me3 in SKBR-3, MDA-MB-231, and MCF-7. These findings provide a framework for understanding the coupling between the chromatin remodelers, chromatin decompaction, and their spatial distribution in non-cancerous and cancerous cell lines. The jacked ratio of euchromatin and heterochromatin in TNBC can be targeted as a potential biomarker for the prediction and prognosis of TNBC.
{kind=link}