In this study involving two groups of 11 healthy humans each, we applied FUS to two brain targets, one cortical (i.e., IFC) and one deep (i.e., Thalamus). Our study is especially unique due to the very high temporal dynamic resolution we used (i.e., one minute). Our findings emphasise that: i) FUS effects are mainly time-constrained and connected to reduced functional connectivity with the parameters used; ii) FUS causes connectivity alterations in the functional networks of the target, relating to the cerebello-thalamo-cortical network following Thalamus-FUS and to multiple networks changes after IFC-FUS; iii) IFC-FUS is directly associated with behavioural alterations through the acoustic pressure applied to the IFC and the IFC-post-central cortex disconnection.
This study was innovative in that we addressed one of the ongoing challenges which prevent the community from fully understanding FUS outcomes: the significance of the delay between the stimulation and its effects. We addressed it with an innovative whole-brain and high time-resolution approach, on healthy humans and two different brain targets. with substantial sample sizes (two groups of 11) and considerable rs-fMRI scan duration (42-minute), particularly notable in human FUS research. As one of the main outcomes, we identified a significant interrelation between the applied acoustic pressure, brain connectivity changes and behavioural alterations. This could lead to a more systematic use of FUS in tailored protocols which aim to change behaviour with a high degree of replicability.
However, some limitations deserve to be mentioned. First, we compared our experimental sessions to our control one, which is a post vs. pre-FUS comparison, without a proper placebo (i.e., sham stimulation). Despite this, our findings remain robust because: i) we used a very stringent high significance threshold (i.e., Family-wise error rate correction) which reduces the possibility of false discovery; and ii) our outcomes are specific to each brain target group, indicating that they arise from FUS rather than participant awareness. Second, we could argue that differences in the parts of the brain reached for each participant may have affected our results (see Fig. 1, panel B, Fig.A1 and Fig.A2). While FUS spatial accuracy surpasses other non-invasive neuromodulation methods, stimulation effects are not restricted to the target area, especially for deep targets. Depending on the used trajectory, the depth of the target and the skull thickness, the consequences of FUS could be different. It is not possible to determine this kind of effect, except through acoustic simulations, which are also mathematical estimations and not real measures. This is, to date, an inherent limitation to using FUS, and requires the development of more accurate devices and methodologies in the future. Third, the rs-fMRI were obtained ~ 15–20 minutes following FUS. It is likely that we missed immediate and short-term effects. Future studies should consider performing online rs-fMRI with FUS to address this issue.
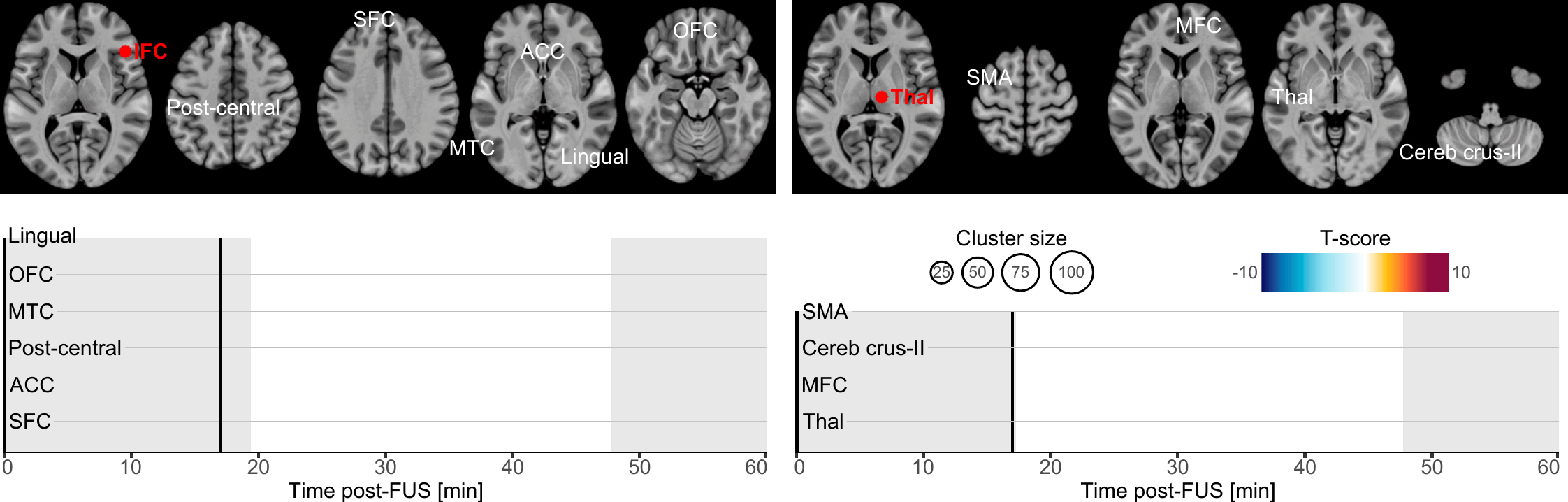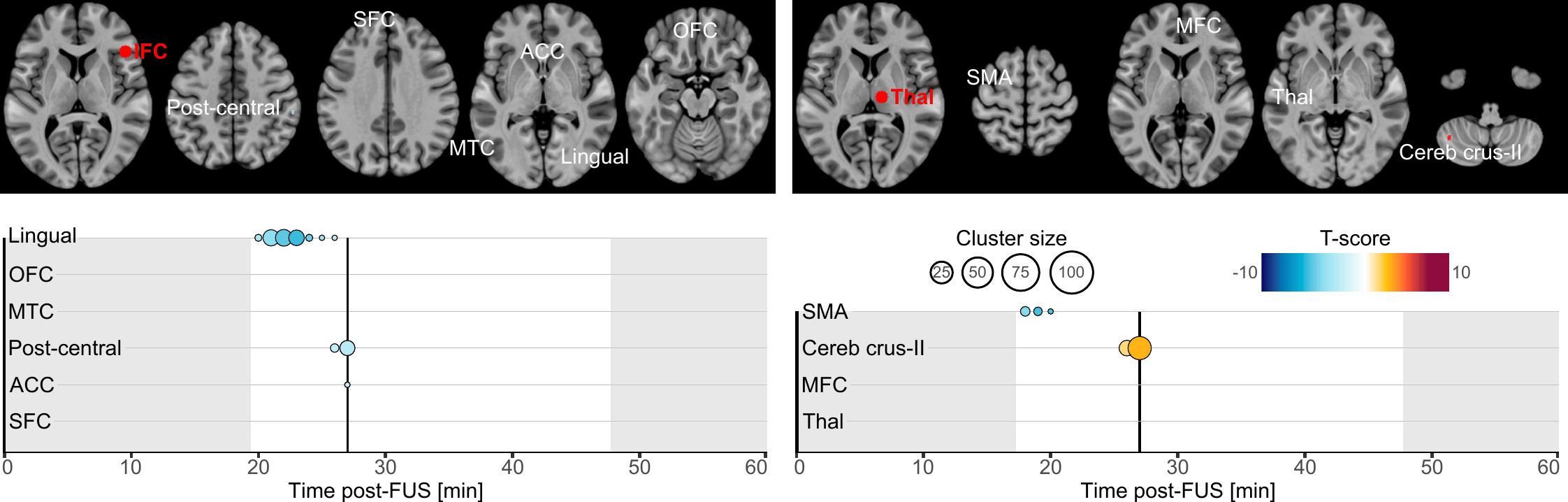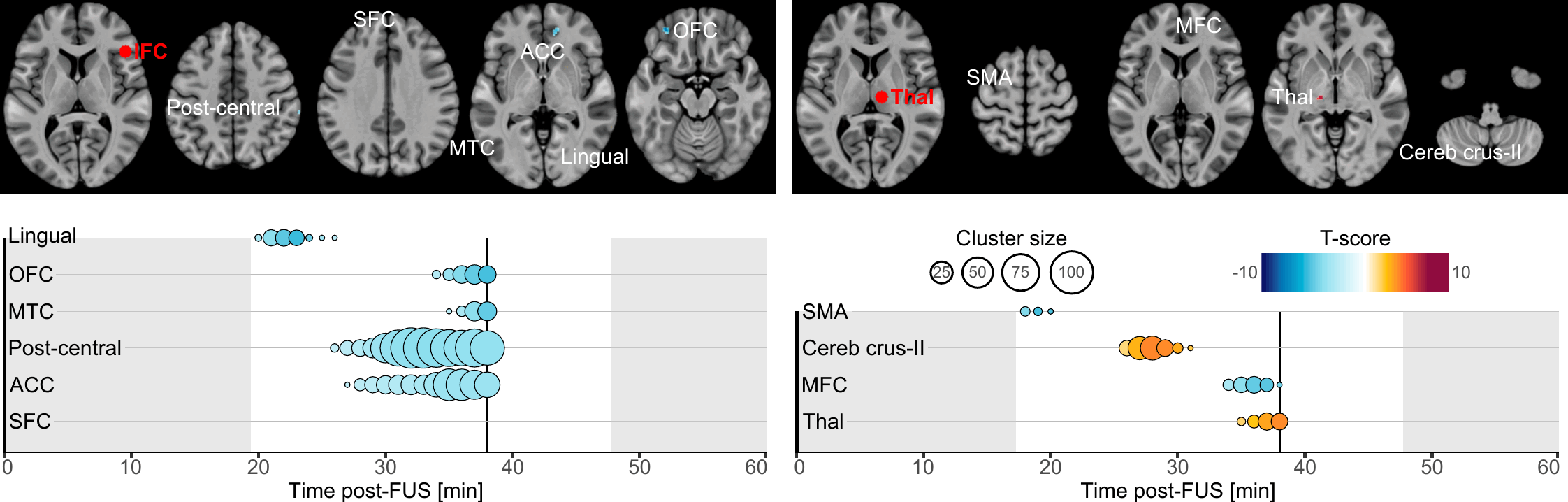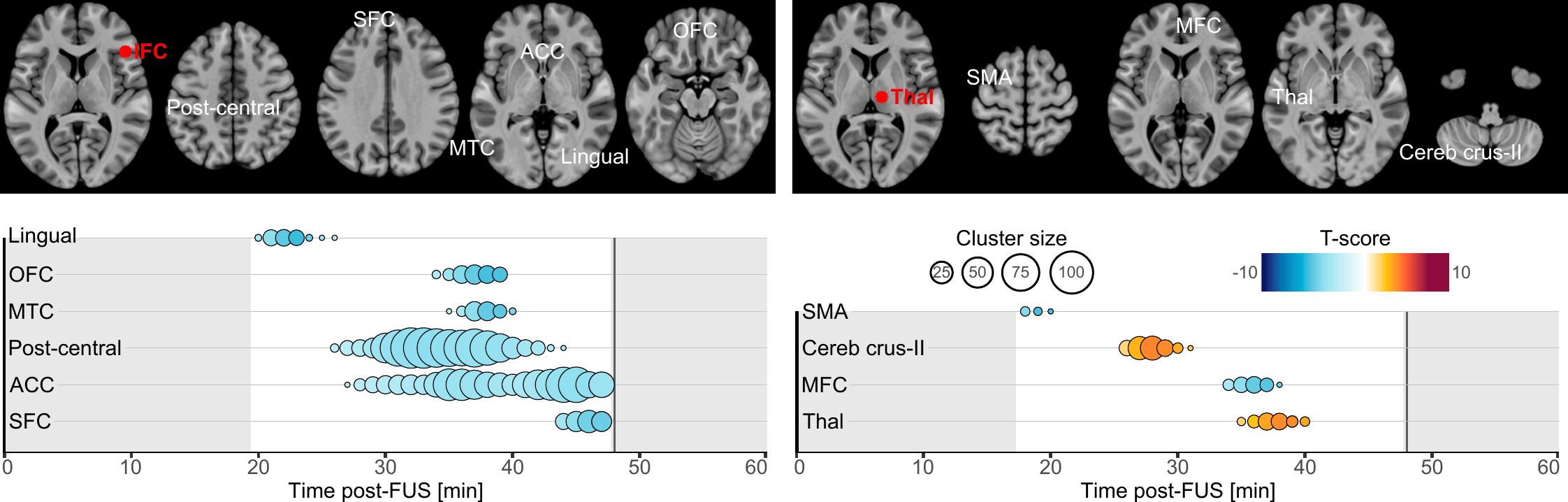
Our primary finding is about the onset and duration during which brain connectivity was altered by FUS. Altogether, our results highlight that brain connectivity was changed in a time-constrained manner which suggests a chain reaction manifesting by a spreading of the FUS effects. Indeed, whilst most of the clusters exhibited transient effects lasting only a few minutes, some persisted near our observation window borders, hinting at potentially longer-lasting effects (lingual cortex, ACC and SFC following IFC-FUS and SMA following Thalamus-FUS), and two were found to have a significantly longer duration (post-central cortex and ACC following IFC-FUS). This underlines the temporary yet significant effect of FUS, and it also reinforces the conclusion that a single 80-second stimulation session is enough to induce changes in brain function for a significant period of time[16]. To some extent, future studies could address the question of how many sessions are needed to induce permanent or long-term changes.
A second significant outcome is the strong relation we observed between FUS effects and the functional connectivity collected during the control condition, but not the structural connectivity (see Fig. 2, panel C). This result is promising in that it means that FUS consequences could reasonably be anticipated to occur within a known brain network. By learning from older neuromodulation approaches such as deep brain stimulation, we know that treatment should be individualised based on various data, including brain connectivity profile[53, 54]. Future studies will have to identify biomarkers related to FUS outcomes and then develop reliable models to maximise clinical improvement while mitigating adverse effects.
As evident in our results, Thalamus-FUS modulates the functioning of the cerebello-thalamo-cortical network in a time-distributed way: decreased connectivity with the left SMA (18–20 minutes post-FUS); increased connectivity with the contralateral cerebellum (Crus-II part; 26–31 minutes post-FUS); increased connectivity with the contralateral thalamus (35–40 minutes post-FUS). This result is of particular importance since the cerebello‑thalamo‑cortical network is known to be implicated in various brain conditions, involving numerous movement disorders[55], schizophrenia[56] and several neurodevelopmental disorders related to motor abnormalities[57]. Since the role of this network is precisely documented, many studies have already used non-invasive neuromodulation to alter its functioning[58, 59]. Recently, a FUS study was conducted on 10 patients suffering from essential tremor[42]. Following Thalamus-FUS, the authors reported a significant clinical improvement in all participants. While this study was mostly a pilot and did not report any neuroimaging results, our findings support it and reinforce the relevance of targeting the thalamus to improve disorders involving the cerebello-thalamo-cortical network.
Regarding IFC-FUS, we exclusively found decreased connectivity, suggesting a functional disconnection of the IFC. This result contributes to the ongoing debate about which parameters induce excitatory or inhibitory effects. On the one hand, our findings are consistent with some articles which suggest that the parameters we used could be related to an inhibitory effect[5, 6]. But on the other hand, our findings negate a recent study which used the same parameters as us, and found an excitatory effect as elicited by a decreased GABA concentration in one of the targets (i.e., posterior cingulate cortex), but not in the second (i.e., dorsal anterior cingulate cortex) and an overall increased functional connectivity[16]. However, as stated by the latter[16], FUS effects could be state or location-dependent.
While half of our connectivity changes are in the anterior part of the brain (i.e., OFC, SFC and ACC), others are more spatially distributed. Based on the known functional connectivity of the IFC, all the clusters we found are in parts of the brain which are known to be related to the IFC[60]. In detail, our results are distributed among functional networks involved in reasoning (i.e., OFC, SFC and ACC clusters), motor execution (i.e., post-central cluster), social cognition (i.e., MTC cluster) and spatial attention (i.e., lingual cluster)[60]. This high variability could be explained by a lack of spatial accuracy regarding the activated volume by FUS (see Fig. 1, panel B).
Regarding the behavioural effects of IFC-FUS, two findings could be reported. On the one hand, our results do not align with our initial behavioural hypothesis, which was to induce an inhibitory process alteration following IFC-FUS (i.e., as measured by the SSRT). This discrepancy may be attributed to our inability to consistently target the specific subregion of the IFC involved in inhibitory processes, namely the posterior-ventral part[61]. Previous studies exploring the impact of FUS on reactive inhibition have yielded mixed outcomes, as inhibitory process changes were not always observed: improved only when IFC-FUS was applied during STOP trials[29]; decreased SSRT following M1-FUS.
On the other hand, we induced a significant reduction in the response time for the GO trials which was explained by the acoustic pressure applied to the IFC and by the functional disconnection between the right IFC and the post-central cortex (see Fig. 3, panels C, D and E). Importantly, the absence of inhibitory changes associated with this speed improvement suggests that participants did not adjust their behaviour (i.e., going faster despite increased risk of failing STOP trials). Furthermore, this effect could not be attributed to a learning effect, as it was not observed following Thalamus-FUS.
One plausible explanation could be that the disconnection of executive and associative networks (involved in inhibition)[62], through IFC-FUS, did not influence reactive inhibition (as measured by the SSRT) but decreased tonic proactive inhibition played by the IFC to the post-central cortex[63], and therefore released the ability to do a motor action faster. This interpretation is consistent with previous observations of improved response speed following IFC-tDCS[64] or M1-FUS[32] and aligns with the established role of the IFC in both reactive and proactive inhibition[63].
{kind=link}