Relative position and size of STN vs. pSTN differ across the anteroposterior (AP) axis
To assess if mRNA profiles originally identified by transcriptomics of the STN and adjacently located pSTN in the mouse are applicable for the study of corresponding structures in the primate STN, the battery of mRNA markers originating from our transcriptomics study (57) was here applied in serial sections covering the AP axis across STN and pSTN in both mouse and macaque, allowing their comparison (Figure 1 and Figure 2). Selected mRNA markers were subsequently analyzed using light-sheet microscopy to validate findings from the section analysis using 3D imaging of the intact mouse brain (Supplementary Videos 1-3).
FISH labeling supported previous conclusions that Pitx2, Vglut2, Htr2c, Kcnab3, Nxph1, Nxph4 mRNAs are ubiquitously distributed across the STN, so called pan-STN markers, and that Pvalb and Col24a1 are locally distributed in the STN in a gradient manner (Pvalb: dorsal-to-ventral; Col24a1: ventral-to-dorsal), confirming the molecular distinction of the STN into three major domains (STNa Pvalb-/Col24a1+; STNb Pvalb+/Col24a1+; STNc Pvalb+/Col24a1-) (57). Further, Tac1 mRNA was confirmed as a selective pSTN marker, Baiap3 as present in pSTN and adjacent hypothalamus but not STN, and Calb2 as present in ventral STN but primarily in pSTN and adjacent hypothalamus (57) (Figure 1a-d).
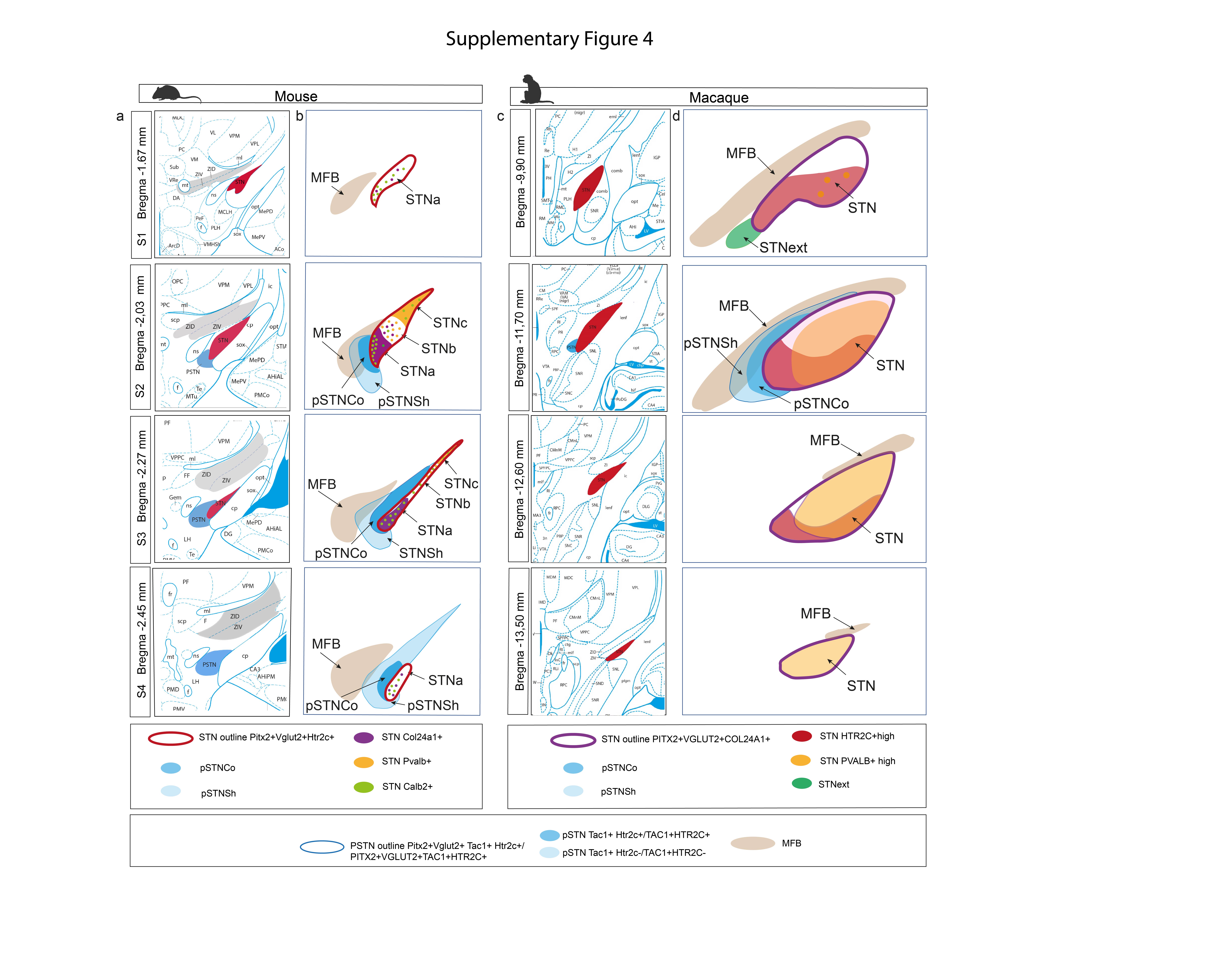
By now comparing multiple combinations of these markers by co-FISH in serial sections throughout the entire area, a striking difference in the overall shape of both STN and pSTN was observed across the AP axis, not recapitulated in published mouse atlases (71) (Figure 1b-d). In serial section FISH analysis at section levels (S) S1-S4, referring to bregma (mm) -1.91 (S1), -2.15 (S2), -2.27 (S3), -2.45 (S4), the anatomical appearance of the STN and pSTN was different in each of the four selected section levels. Thus, while preserving their relative distribution throughout the STN structure, the molecularly defined internal STN domains occupied different amount of space across S1-S4 (Figure 1c,d, with illustrations summarizing the results to the right). At S1 (anterior), only the molecular profile of STNa was seen; at S2, the STN shape was the classical almond shape commonly used to visualize the STN in literature (STNa, STNb, STNc); at S3, the STN was a narrow slice but still positive for markers allowing the subdivision into the three domains; at S4 (posterior), again, only the molecular profile of STNa was seen (Figure 1c,d).
Also the pSTN showed an unexpected spatial distribution. Tac1 mRNA compared with Calb2 and Baiap3 as well as the STN markers clearly showed that the pSTN covers substantially larger space than previously described (Figure 1c,d). In fact, the roundish shape at the medial tip of the STN most commonly used in publications to denote pSTN (Figure 1b) was present at S2 only, while anteriorly (S1), no pSTN was detected, and posteriorly (S3, S4), the shape and spatial relationship STN vs. pSTN was dramatically different. At S3, the pSTN was observed as a narrow but elongated structure, which was aligned with the entire dorsal aspect of the STN (aligning STNa, STNb, STNc). Thus, at S3, analysis of several markers clearly showed that the cellular densities of STN and pSTN together form a shape which is very similar to the characteristic STN shape observed at S2 (and illustrated in most atlases, (71)) (Figure 1c,d). Finally, at S4, the most posterior position of the STN (STNa), the pSTN is at its largest and wraps around the STN, with an outline that resembles the almond-like STN structure at S2 (Figure 1c,d). Further, not only STN but also pSTN showed molecularly heterogeneity; rare Pitx2-positive cells were observed in pSTN which all co-localized with Vglut2, but not all Vglut2-positive pSTN cells were positive for Pitx2; further, Tac1 and Calb2 mRNAs did not show complete overlap, suggesting they represent different pSTN subpopulations (Figure 1a-d).
These molecularly enhanced anatomical details were firmly confirmed using 3D light-sheet imaging of the intact mouse brain, applying fluorescent probes directed at Pitx2 and Tac1 to visualize the spatial distribution of STN and pSTN, respectively. In coronal fly-through (set at slower speed at the level of STN and pSTN to clearly outline the spatial distribution of the markers), Pitx2(Supplementary Video 1), Tac1 (Supplementary Video 2), and Pitx2/Tac1 combined (Supplementary Video 3) allowed the entire STN/pSTN complex be spatially revealed within the brain, outlining the structural relationship between STN and pSTN across all axes.
In conclusion, mRNA markers enabled the visualization of anatomical features that have not previously been described for the mouse brain, such as a parallel alignment of STN and pSTN as pSTN gradually increases in size relative to STN posteriorly.
Ventromedial STN defined by a distinct Serotonin receptor 2c profile in macaque
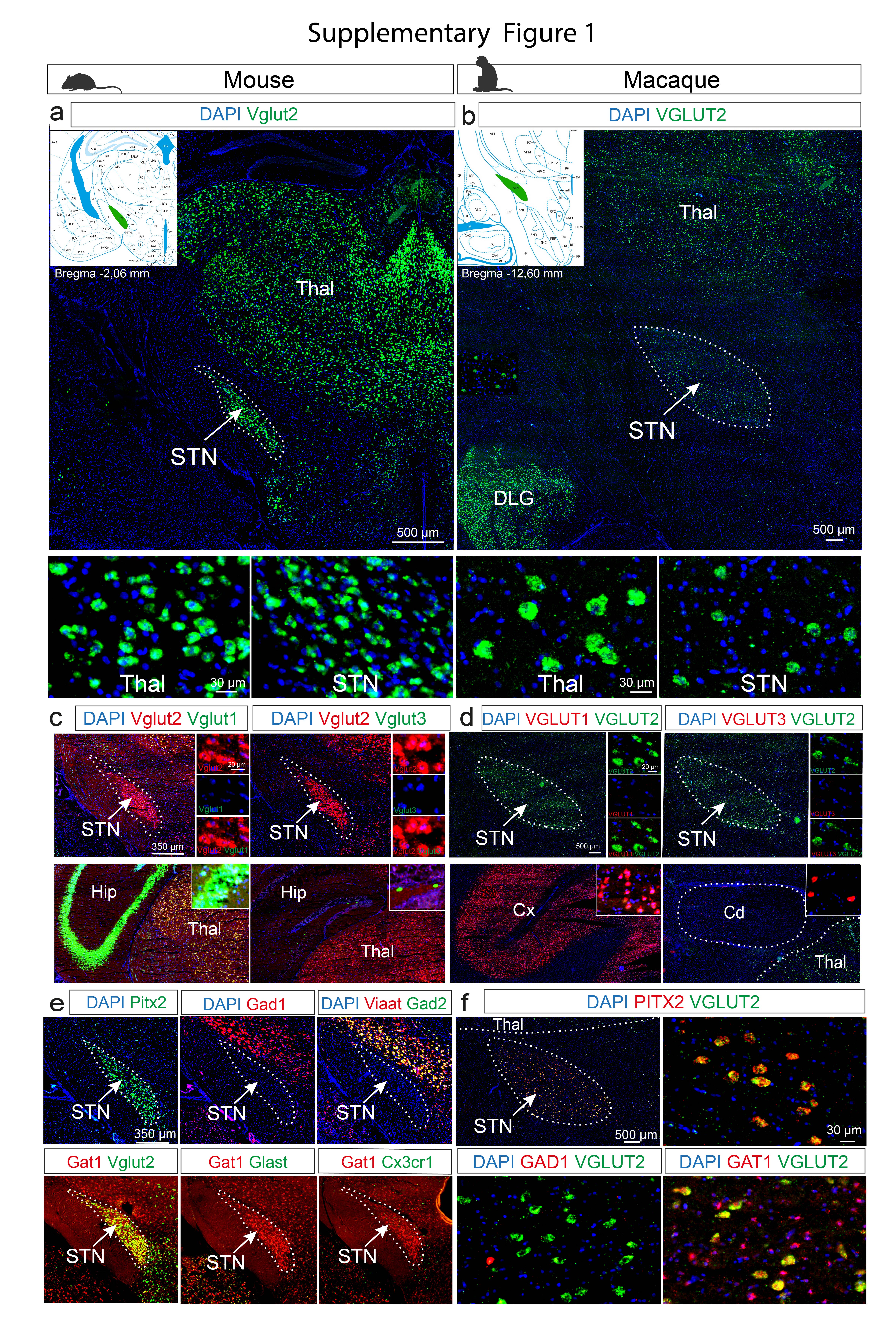
The macaque brain was subsequently analyzed. First, the question of neurotransmitter identity was addressed. The STN is well known as primarily excitatory in nature (72–76), however, the presence of inhibitory neurons and glia cells has remained less explored. By co-FISH analysis, the vast majority of STN cells were identified as positive for the glutamatergic marker Vglut2 in both mouse and macaque, while Vglut1 and Vglut3 were not detected, and only rare cells were positive for inhibitory markers Gad1, Gad2, and, Viaat mRNAs. Similar to mouse STN, the macaque STN (mRNAs in capital lettering) was also positive for PITX2, overlapping completely with VGLUT2. In contrast to rare inhibitory markers, almost all STN neurons in mouse and macaque were positive for Gat1 mRNA, a marker for cells with capacity for transport of extracellular GABA (77). Finally, glial markers Eaat1/Glast1 or Cx3cr1 mRNAs were detected in only few cells mouse and macaque STN (Supplementary Figure 1). Thus, the strong glutamatergic phenotype of the STN was verified in both mouse and monkey, with indication of sparse presence of inhibitory neurons and glia cells. Further, PITX2, the primate counterpart of Pitx2, a well-described STN marker in mice (61,62,67), was identified as marker for VGLUT2+ neurons throughout the STN also in the macaque.
Next, co-FISH was performed, analyzing the differentially expressed genes originating from snRNASeq in mice (57). Serial section across the entire STN area in the macaque brain identified NXPH4, NXPH1, KCNAB3 mRNAs as pan-STN markers, present across the STN in a similar manner to PITX2, and also in this species, co-localizing 100% with VGLUT2 (Figure 2a,b). The pSTN and several adjacent structures were positive for BAIAP3, a marker excluded from the STN in both species (Figure 2a,b). The pSTN in macaque was also positive for TAC1, as recently described (28); further analyzed below.
Apart from these similarities between mouse and macaque, one striking difference was detected. COL24A1 mRNA, in the mouse a distinct marker for the ventromedial STN (shown in Figure 1), was present throughout the whole STN structure in the macaque in a ubiquitous manner, overlapping with VGLUT2 (Figure 2c). In contrast, HTR2C mRNA, encoding the serotonin receptor subtype 2c protein (5-HTR2c), which in the mouse was ubiquitously present throughout the STN, was restricted to the ventromedial aspect of the STN in the macaque (Figure 2c). Thus, the ventromedial STN is defined by Col24a1 mRNA in mouse, but by HTR2C mRNA in macaque; these two mRNA showing differential localization in the STN of mouse vs. macaque (Figure 2b-d).
Serotonin receptor 2c mRNA defines a ventromedial domain in macaque STN particularly prominent in its anterior aspect
The striking pattern of Htr2C mRNA in the ventromedial STN of the macaque was of particular interest to address further, not least from the perspective of the importance of the serotonergic system in pharmacology-based medical treatments of neuropsychiatric disorders, including OCD and depression. Indeed, serotonergic innervation of the STN structure has been shown in several studies (19,40,41,78–80), including a recent study in which we identified serotonergic innervation selectively in the anteromedial STN (60).
In the primate (monkey and human) and rat, the parvalbumin protein (PV) has been shown in several studies as a marker of dorsal and lateral STN (77,81,82), and we recently described a similar distribution of its corresponding mRNA (PVALB) in macaque (28). To now investigate if PVALB and HTR2C mRNAs form overlapping or discrete profiles throughout the STN, the distribution pattern of Htr2C mRNA was next compared to PVALB in the monkey (Figure 3).
At section levels S1-S4 (bregma (mm) -10.35 (S1), -11.70 (S2), -12.60 (S3), -13.50 (S4), with S1 representing the anterior STN), PVALB and HTR2C mRNAs showed both overlapping and distinct distribution, depending on section level (Figure 3a,b). PVALB mRNA was the most dominant of the two mRNAs but both showed regional distribution across the STN. Anteriorly (S1), only a small medial STN area was positive for PVALB, while posteriorly (S2-S4), the PVALB+ area covered the majority (but not all) of the STN. In contrast, HTR2C mRNA was most prominent anteriorly (S1). HTR2C covered more than half of the STN at S1 and was most prominent in the ventromedial STN. HTR2C decreased posteriorly, but was always found located in a ventromedial strip of the STN (S2-S4), positive in the medial tip throughout the STN (Figure 3a,b). HTR2C/PVALB overlap was observed in ventral STN but never in the medial tip, due to the fact that PVALB mRNA was consistently excluded from this STN area in which HTR2C was exclusive at all levels (Figure 3a). PVALB was more dominant posteriorly, while HTR2C dominated anteriorly (Figure 3a,b). Thus, apart from limited overlap, PVALB and HTR2C were mutually exclusive in the STN, and both mRNAs showed rather sharp borders (Figure 3a-c, close-ups of borders).
The intensity level of the VGLUT2 FISH signal in the macaque brain was higher medially and ventromedially than laterally (Figure 3e).However, both PVALB and HTR2C overlapped extensively with VGLUT2 mRNA, within their regional distribution areas (Figure 3e). Using VGLUT2 as pan-STN glutamatergic marker, quantification of cells positive for PVALB vs. VGLUT2, or HTR2C vs. VGLUT2, and direct comparison PVALB vs. HTR2C confirmed the observation by visual inspection above of more abundant HTR2C than PVALB labeling anteriorly and ventromedially, and vice versa posteriorly and laterally. In a total STN count (not taking precise position into account), PVALB+/HTR2C- was the dominating combination of STN neurons (77%), with similar amounts of cells positive for both PVALB and HTR2C, or selective for HTR2C (PVALB+/HTR2C+, 13%; and PVALB-/HTR2C+, 10%) (Figure 3d). Next, to assess if these patterns are reproduced in the human STN, biopsy sections were addressed, revealing a similar distribution of PVALB and HTR2C mRNA in human STN as in the macaque STN (Figure 3f,g).
Concluding these findings, along the axes of the macaque and human STN, three domains can be identified, each reflecting the spatial distribution of PVALB and HTR2C: STNPVALB+/HTR2C-, STNPVALB+/HTR2C+, STNPVALB-/HTR2C+. The STNPVALB-/HTR2C+ molecular phenotype is most abundant in the ventromedial STN, and includes the medial STN tip while also dominating in the anterior STN (illustrated in Figure 3c,g).
Unique molecular profile identifies a far-anterior STN domain in the primate
While addressing the spatial distribution of HTR2C in macaque STN, positive cells were observed anterior to the area outlined in atlases as the STN. This was curious and called for more careful assessment. Thus, to further investigate macaque brain anatomy, the proximity of the posterior STN to substantia nigra (SN), and of the anterior STN to hypothalamic structures serial sections were analyzed by histological cresyl violet staining (Figure 4a) and FISH for TH, VGLUT2 AND GAD1 (Figure 4b,c). At the AP level of posterior hypothalamus, a cellular density was found in connection with STN, but which does not appear described in primate brain atlases (Figure 4a-c). To analyze if this might be an overlooked STN or pSTN domain, the molecular profile of this density was analyzed using the same battery of STN and pSTN mRNA markers as implemented above.
First, VGLUT2/GAD1 co-localization analysis showed that this anterior density of cells was enriched in both VGLUT2 and GAD1, but not their overlap. More cells were positive for VGLUT2 than GAD11, but quantification revealed a stronger GAD1-presence in this far-anterior cellular density than in the regular STN (35% vs. 1% GAD1-positive cells in far-anterior vs. regular STN; 7% in pSTN) (Figure 4d). This far-anterior aspect, rich in both VGLUT2 and GAD1, was given the working nick-name vgrR (VGLUT2/GAD1-rich Region) (Figure 4b-d).
Next, to determine if molecular profiles could help determine if this far-anterior structure was of STN or pSTN identity, further mRNA analysis was performed (Figure 4e-g). Results showed that vgrR was negative for PVALB, the marker of dorsal STN, and positive for HTR2C, the marker of anterior and medial STN, described above. Further, vgrR was positive for macaque pan-STN markers PITX2, KCNAB3, and NXPH4 (which all co-localised to 100% with VGLUT2), while completely negative for the pSTN marker TAC1, and almost completely negative for CALB2 and BAIAP3 (<5%), the markers most positive in pSTN and posterior hypothalamus (Figure 4e-g).
In summary, apart from GAD1-positive cells, the vgrR area shared its molecular phenotype with that of the anterior STN, including the PVALB-/HTR2C+ profile. However, the combined VGLUT2/GAD1 positive molecular phenotype distinguished vgrR from the remaining STN. This mixed neurotransmitter phenotype might reflect the location far-anteriorly, and based on its molecular profile, vgrR will here be referred to as the “far-anterior extension of the STN” (STNext) (illustration in Figure 4h). Notably, STNext could not be detected in the mouse, and is thus a primate feature.
Serotonin receptor 2c mRNA allows distinction of pSTN into two domains: pSTN core and pSTN shell
In light of the striking localized pattern of HTR2C mRNA in the ventromedial STN of macaque and human brains, and given the possibility to use this marker to help define the previously undescribed STNext area of the macaque brain, the presence and putative distribution of HTR2C mRNA was next explored in the pSTN, revealed above as a more spatious structure than recognized, and to partially align the posterior STN in mice (Figure 1). Similar to the STN, serotonergic innervation of the pSTN has been shown (83,84). Now, it was of interest to detect if pSTN might be selectively positive for HTR2C in macaque, in a manner similar as revealed above for STN (Figure 2,3).
HTR2C was next assessed in the pSTN by comparison with the pSTN marker TAC1, originally described as marker for distinct pSTN neurons in mice (57) and recently identified as selective for a subpopulation of pSTN neurons also in macaque (28). HTR2C and TAC1 mRNAs were analyzed across the STN-pSTN area in mouse (Figure 5a,b) and macaque (Figure 5c,d).
First applying co-FISH analysis across S1-S4 levels in the mouse, and paying attention to the structural differences of STN and pSTN across the AP axis described in Figure 1, Htrc2 mRNA was confirmed as strongly positive across STN while pSTN was selectively positive for Tac1 (Figure 5a,b). However, Htrc2 and Tac1 mRNAs were not mutually exclusive. Instead, Htr2c was could be detected also in pSTN, albeit at much lower detection levels than in the STN (Figure 5a,b). Tac1+/Htr2c+ double-positive pSTN cells (with moderate Htr2c labeling compared to STN cells) were found primarily in a domain bordering to the STN, while Tac1+/Htr2c- cells (low or zero Htr2c labeling) were identified more laterally. This difference in Htr2c between medial and lateral pSTN allowed the subdivision of pSTN into a core (Co) and shell (Sh) area: pSTNCo (pSTNCo;Tac1+/Htr2cmedium) closest to the STN, and pSTNSh (pSTNSh; Tac1+/Htr2cweak), lining the pSTN core laterally (Figure 5a,b). In contrast to the sharp borders of Tac1 and Htr2c, Calb2 was detected throughout pSTN (Figure 5b).
Across the AP axis in the macaque, and by using VGLUT2 as reference, TAC1 mRNA was identified in pSTN at section levels S1 and S2, but not S3 and S4; macaque pSTN thereby lining the STN anteriorly, but not posteriorly. At S1, TAC1 cells were observed close to the anterior aspect of the STN (medial STN tip and STNext), while at S2, TAC1-positive cells covered a substantially larger area lining the dorsal STN (Figure 5c). Analysis of HTR2C and TAC1 thus demonstrated that the macaque pSTN covers a major area directly associated with anterior STN. Further, HTR2c allowed the subdivision of pSTN into a core and shell area in similar manner as for mouse, and many pSTN cells positive for Calb2 mRNA also in macaque (Figure 5d).
Concluding, based on mRNA profiles of TAC1 and HTR2C, the pSTN is anatomically associated with anterior STN in the primate, and with posterior STN in the mouse. In both species, the differential levels of serotonin receptor 2c mRNA allows the subdivision of pSTN into a core and shell area (summarized in Figure 5e).
pSTN, but not STN, cells embedded in dopaminergic nerve bundles of MFB in both mouse and macaque.
At its dorsal aspect, the STN borders to the monoaminergic projections of the MBF. With the present observation that pSTN aligns with the dorsal STN, it was of interest to address if pSTN and MFB co-localize above the STN. The MFB, which is a projection bundle, has been shown to contain certain cell groups of various identity, including hypothalamic cells (20,21), however, pSTN cells have not previously been associated with the MFB.
To now address if these three aligning brain structures (STN, pSTN, MFB) were overlapping such that STN or pSTN cells are embedded in the MFB, molecular profiles were addressed in co-localization analysis on serial macaque sections using TH immunoreactivity as a marker for the dopaminergic identity of the MFB. Indeed, TH-positive projections within the MFB align with both the STN and pSTN, including the far-anterior STNext (Figure 6a). TH-positive fibers contained ball-like globular structures, positive for TH, and DAPI staining showed presence of cell nuclei in the MFB (Figure 6b,c). In combined immunofluorescent/FISH analysis addressing TH protein and either VGLUT2 or TAC1 mRNAs (Figure 6b, TH/VGLUT2; Figure 6c, TH/TAC1), MFB (TH+) was seen to directly border the dorsal aspect of the STN and pSTN. Further, VGLUT2+/TAC1+ cells were discovered within the TH-positive fiber bundles (Figure 6b,c; illustration in Figure 6d).
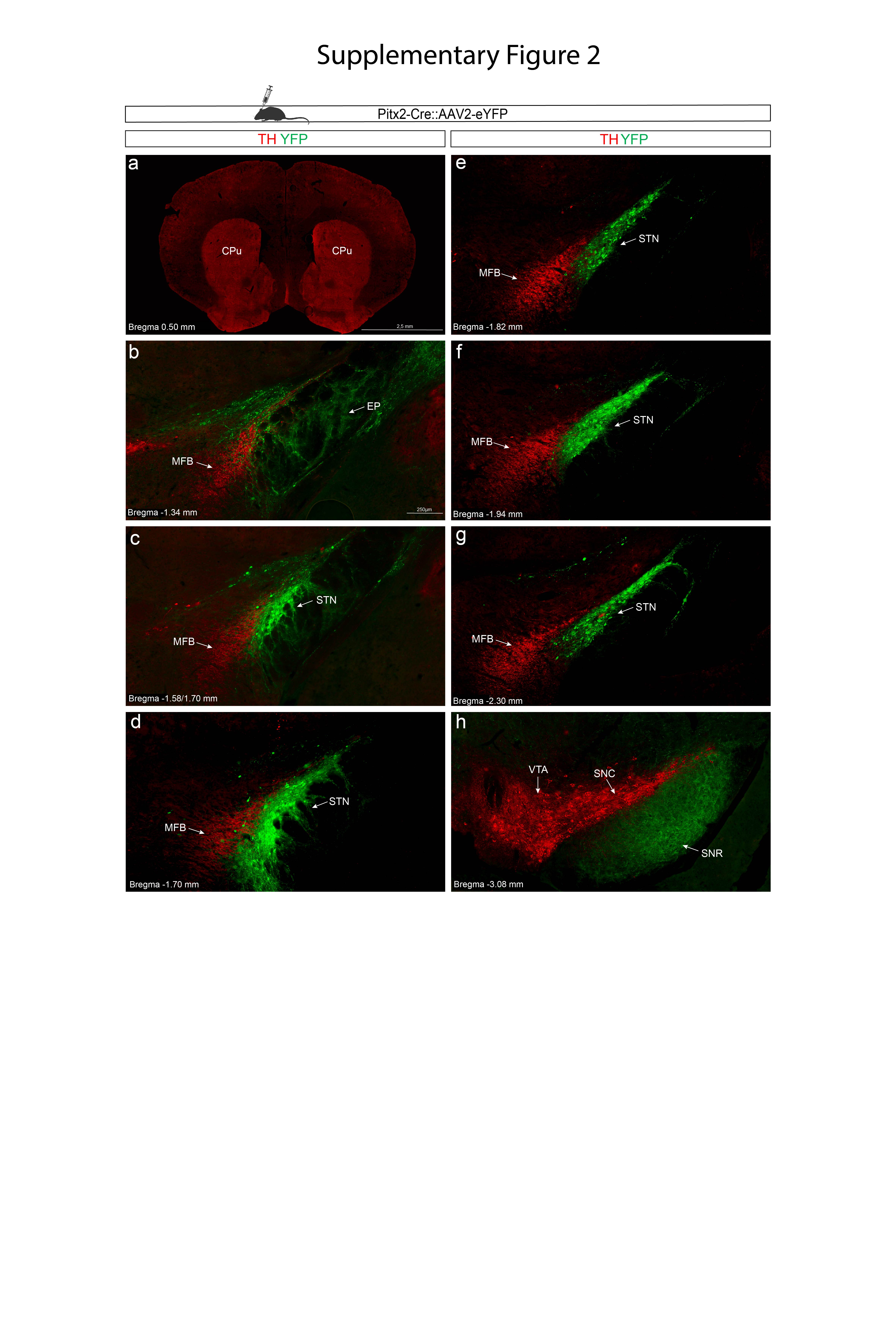
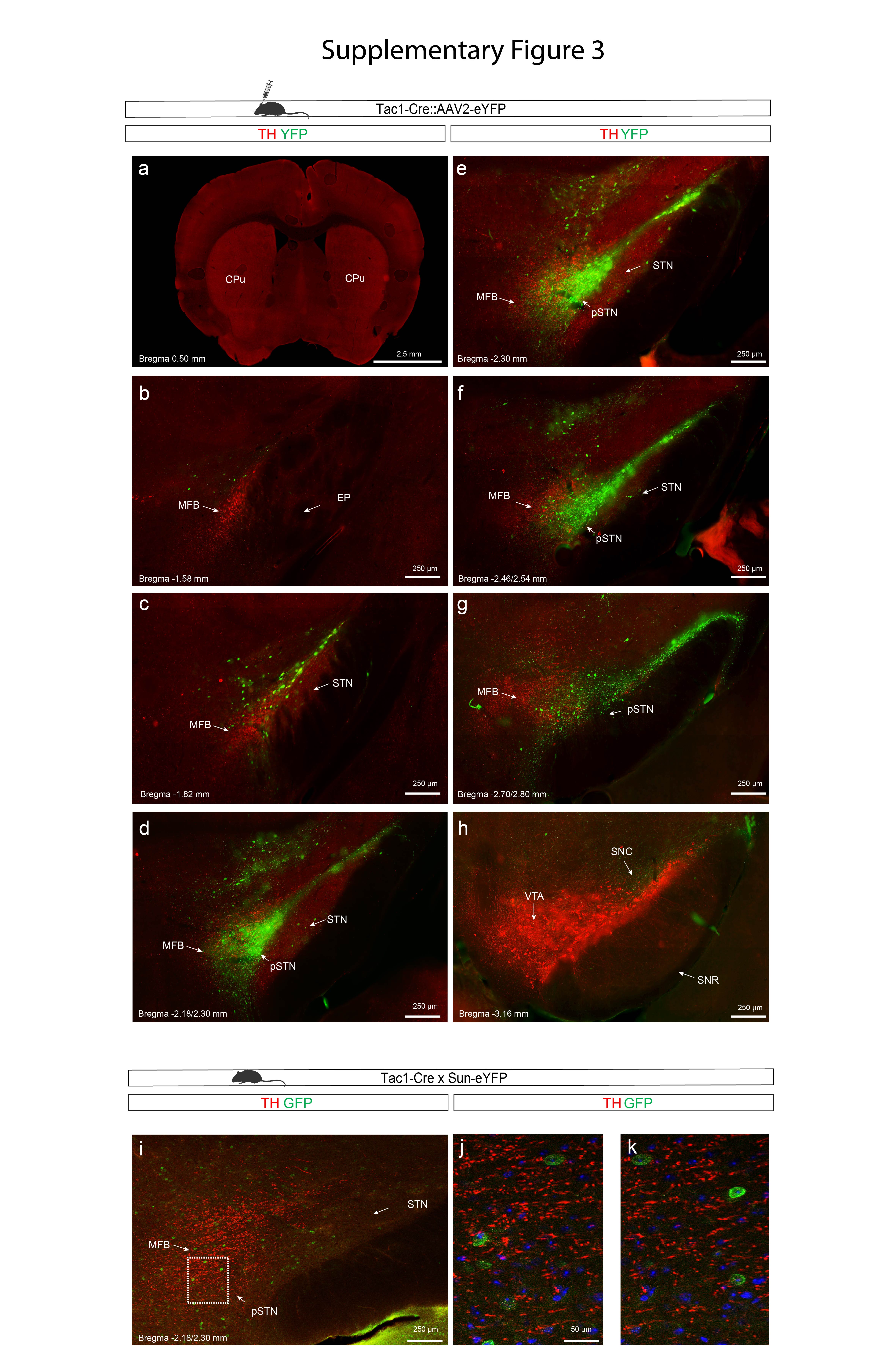
To further assess the observation of TAC1+ neurons embedded within the MFB, and exclude that this cellular feature was a tissue-processing anomaly, cellular tracing using viral-genetics methodology in recombinase-expressing mice was next implemented. In contrast to the monkey, no globular ball-like TH-positive structures were observed in the mouse MFB (Figure 6e,f). However, upon injection of adeno-associated virus (AAV) containing a double-floxed reporter gene construct encoding the Yellow fluorescent protein (YFP) into the STN and pSTN of Pitx2-Cre and Tac1-Cre transgenic mice, and comparison of YFP-positive cells with TH immunofluorescence, YFP-positive cells were amply detected within the mouse MFB. Importantly, this was only observed in the Tac1-Cre mouse line, reflecting pSTN neurons, and not in the Pitx2-Cre mouse line, showing STN neurons. Thus, pSTN, but not STN, neurons are embedded in the MFB in both mouse and primate (Figure 6e,f; Supplementary Figures 2,3).
Finally, using light sheet-microscopy for whole mouse brain 3D imaging analyzing Pitx2 (STN) combined with TH immunofluorescence (Supplementary Video 5) and Tac1 (pSTN) combined with TH immunofluorescence (Supplementary Video 6), coronal flythrough allowed the visualization the of STN and pSTN distribution along the TH-positive MFB, and, while weakly detected, also validated the presence of Tac1+, but not Pitx2+, cells within the MFB at the dorsal aspect of STN.
{kind=link}
{kind=link}
{kind=link}
{kind=link}