4.1. Search results
A total of 1372 articles were collected using Scopus (n = 632), Web of Science (n = 469) and PubMed (n = 271), with 307 duplicates removed. Following title and abstract screening, 948 irrelevant articles were removed. 117 full-text articles were assessed for inclusion, with 28 excluded (Figure 1). Three of these were excluded due to use of an inappropriate model 30–32, two for being performed in vitro 33,34, two due to not being relevant to TGF-β1 35,36, eight for not having a full text available 37–44, one due to a lack of experimental controls 45 and 12 for measuring inappropriate outcomes or measuring TGF-β1 as part of a large panel of molecules 46–57. As such, 89 studies were included for analysis in the present review.
4.2. Modelling of stroke in pre-clinical animal studies: study design
4.2.1. Ischemic stroke models
Some studies reviewed used multiple methods or animal species within the same body of work, which are counted separately below. Of the 89 included studies (Table 1), 68 modelled ischemic strokes, with middle cerebral artery occlusion (MCAO) by insertion of an intraluminal filament being the most widely used model. Of these, 52 studies employed the transient MCAO (tMCAO) or MCAO/reperfusion (MCAO/R) model, whilst 12 utilized a permanent MCAO approach, typically with clamps or cauterization. A further four studies used photothrombosis to induce focal cortical ischemia following administration of a photosensitizing agent, whilst two studies utilized the injection of autologous blood clots to the internal carotid artery.
MCA occlusion time was highly variable within tMCAO studies, with a range between 20 minutes to 2 days of occlusion prior to reperfusion, as is depicted in Figure 2. 60, 90 and 120 minutes were the most common occlusion times, collectively accounting for 36 of the 89 included studies.
4.2.2. Hemorrhagic stroke models
Of the 89 included studies, 21 modelled hemorrhagic stroke (Table 2); 8 SAH, 7 ICH and 6 GMH/IVH. SAH was induced by endovascular perforation of the middle cerebral artery (MCA) or intracranial internal carotid artery (ICA), or by injection of blood/blood products into the cisterna magna. ICH was induced by injection of blood products or collagenase into the brain parenchyma, and IVH/GMH was induced by intracerebroventricular injection of blood, collagenase or glycerol.
4.2.3. Animal species and strains used
Rodent models were used in nearly all included studies for both forms of stroke (Figure 3). Of the 89 included studies, 53 used rats; 33 studies used Sprague-Dawley (SD) rats, 14 studied Wistar rats, 3 used spontaneously hypertensive rats (SHR), one used Lewis rats and one study did not disclose the rat species used. Of the 31 studies which used mice, 19 used the C57BL/6 or C57Bl/6J strains, four used the CD-1 strain, one used Kunming mice and one the BALB/C strain. Seven studies utilized transgenic mice, and two studies did not disclose the mouse strain. Additionally, two studies used white New Zealand rabbits and one used piglets. Only two studies utilized non-human primates, with one using rhesus monkeys and one using baboons.
Male animals were used in 73 of the included 89 studies (81.8%), with only eight studies using mixed sex populations and five using only female animals (Figure 3, inset). In addition, three studies failed to disclose the sex of the animals employed within the study.
4.3. TGF-β1 measurements
TGF-β1 was measured directly in 52 ischemic stroke studies (76%) and 13 hemorrhagic stroke studies (62%). In ischemic stroke the main measures were semi-quantitative, employing PCR (16 studies) and in situ hybridization (4 studies) to assess gene expression, and western blot (26 studies) and immunohistochemistry (20 studies) for assessing protein in a variety of samples; whole brain, contralateral hemisphere, ipsilateral hemisphere, penumbra, core and peri-infarct cortex. Assessments were usually done within the first few days of stroke induction, with only a few studies conducting time course experiments. One study 58 demonstrated TGF-β1 gene expression peaked at 7 days post MCAO in the core, had increased slightly in the peri-infarct area but remained low in the contralateral hemisphere, showing different patterns of expression in the different areas of the lesion. Only 6 studies used enzyme-linked immunosorbent assays (ELISA) or Luminex to get quantitative TGF-β1 levels, mainly demonstrating TGF-β1 increased in brain tissue following ischemic stroke, although the values were extremely variable (186.2 to 53,200 pg/g in MCAO, as shown in Additional File 1, Table 4).
In hemorrhagic stroke, western blot (7 studies) and ELISA (6 studies) were the most frequent techniques for measuring TGF-β1, followed by PCR (4 studies) and immunohistochemistry (3 studies). TGF-β1 was mainly measured by ELISA in CSF from IVH or SAH studies and demonstrated an increase in the levels of TGF-β1 protein over 2-3 weeks 59–62 with very consistent concentrations (maximum of 69.3-315.5 pg/ml). One study in rabbits did demonstrate a reduction in CSF TGF-β1 following IVH but their control levels were variable 63.
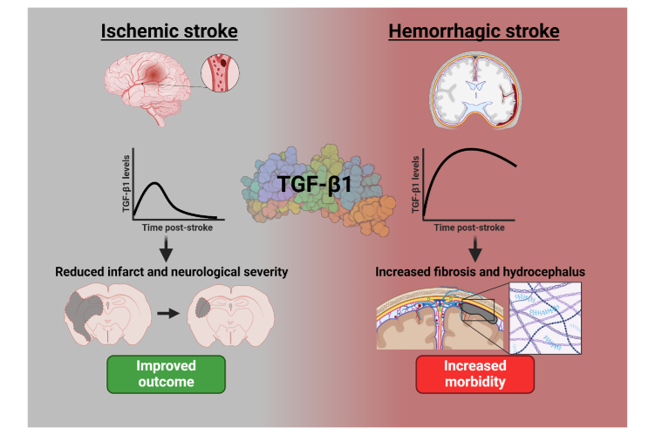
4.4. Protective or harmful role of TGF-β1
Preclinical models of ischemic and hemorrhagic stroke showed a split in the possible protective role of TGF-β1, as depicted in Figure 4. Fifty-two of the included 68 ischemic stroke studies (76%) showed a protective effect of TGF-β1 supplementation or expression, with 10 (15%) showing a non-protective effect and 6 (9%) showing an unclear effect. Within hemorrhagic stroke studies, 7 of the 21 included studies (33%) showed a protective effect of TGF-β1, 11 (57%) showed a non-protective effect and 3 were unclear (14%).
4.5. TGF-β1 in pre-clinical ischemic stroke studies
Fourteen ischemic stroke studies were performed with no intervention, typically examining TGF-β1 protein or RNA concentrations in brain tissue or exploring the role of components of the TGFβ pathway in functional recovery. An increase in TGF-β1 expression was noted specifically in the cortex 64,65, ischemic core 66, cervical spinal cord 67 and hypometabolic zones 68 post-MCAO. An increase in TGF-β1 levels or activation was shown to improve the number of NG2+ microglia 58 and CD68+ microglia/macrophages 69, increase neuroprotection 70, angiogenesis 20 and vascular remodeling 71 and correlated with spontaneous recovery 72–74. In addition, suppression of the TGF-β1/Smad3 pathway by miR-323 was shown to increase apoptosis post-stroke 75.
Only 24 ischemic stroke studies (36%) modulated TGF-β1 directly, either by giving exogenous TGF-β1/overexpressing TGF-β1 (13 studies) or inhibiting TGF-β1/receptor (14 studies). Exogenous TGF-β1 was commonly used as an intervention and shown to reduce hemorrhagic transformation (HT) (2.5 µg IV) 76, limit neuronal injury (50 ng intracerebroventricular (ICV)) 77 and reduce infarct volume and motor deficit (0.25 ng intracisternal) 78. TGF-β1 was also shown to reduce infarct volume, apoptosis and improve neurological function (1 µg intranasal) 79, reduce infarct volume and improve CBF (10/50 µg, internal carotid injection) 80 and improve neurological outcome (1 ng/g) 81 post-stroke. Hemorrhagic transformation (HT) was reduced by exogenous TGF-β1 in monocyte/macrophage depleted mouse brains 82, and by the tyrosine kinase inhibitor Imatinib, which increased TGF-β1 expression 83.
One study demonstrated the anti-inflammatory effect of TGF-β1 (50/100 ng cortical injection) in suppressing NF-κβ activation but did not examine any functional outcomes 22. No included studies demonstrated worse functional outcomes following treatment with exogenous TGF-β1.
A range of anti-inflammatory agents were employed across the cohort; propolis 84, minocycline 85 and wogonin 86 were shown to increase TGF-β1 levels post-stroke, correlating with improved neurological outcomes or infarct volume. ICV injection of recombinant interleukin-4 activated TGF-β1 signaling, reducing apoptosis, infarct volume and decreasing neurological injury 87. However, one embolic stroke study demonstrated treatment with an anti-inflammatory peptide suppressed TGF-β1 levels which improved neurological outcome 88.
Genetic modification of the TGF-β1/Smad pathway was also used as an intervention. Overexpression of microglial transcription factor zinc finger E-box binding homeobox 1 (ZEB1) was shown to reduce infarct volume and improve BBB functionality in a TGF-β1-dependent manner 89. Smad3 overexpression reduced apoptosis following MCAO 90, whilst TGF-β1 overexpression reduced infarct volume 91 and expression of the pro-apoptosis protein Bad 92. Similarly, overexpression of ALK5, a TGF-β1-family receptor, was shown to improve neurological scoring post-stroke 93. Transgenic inhibition of astrocyte TGF-β1 expression generated increased cortical inflammation and infarct size, in addition to worsened neurological severity 94. However, modification of the TGFβ signaling pathway did show a correlation between increased TGF-β1 expression and worsened outcomes in some studies. Overexpression of leucine-rich α-2 glycoprotein 1 was noted to upregulate ALK1, a TGFβ-family receptor, which correlated with worsened infarct size and neurological deficits in a one-hour tMCAO model 95. Similarly, knockdown of long non-coding RNAs MEG3 96 and RMST 97 was shown to reduce the expression of TGF-β1 and key pro-inflammatory cytokines, correlating with improved outcomes. Adenoviral knockdown of BRD4 modified TGF-β1 signaling, reducing phosphorylation of Smad2/3, resulting in improved infarct size and reduced fibrosis 98.
Antagonistic inhibition of TGF-β1 receptors was used in several studies, revealing TGF-β1-dependent reductions in infarct volume following ischemic stroke 8,99,100. Inhibiting TGFβRII was shown to result in increased HT in a neonatal MCAO model 101. However, one study showed a link between ALK5 inhibition and reduced reactive oxygen species (ROS) damage, correlated with improved outcomes post-MCAO. In addition, one study showed a correlation between TGF-β1 expression and increased fibrosis and reduced perivascular CSF flow in the sub-acute stage of the injury (7-14 DPI) 102.
Post-injury conditioning was investigated with a range of stimuli. Isoflurane post-conditioning (for 60 minutes immediately post-MCAO) was shown to reduce neurological severity score (NSS), infarct volume and neuronal survival or density in a TGF-β1-dependent manner 103–105. Remote ischemic post-conditioning generated an increase in TGF-β1 expression and phosphorylation of Smad2/3 which correlated with reduced infarct volume and improved NSS, confirmed by TGF-β1 knockdown 106. Hypoxic post-conditioning, in which animals were contained in a 5% O2 atmosphere for 45 minutes, was shown to increase ALK5 expression, correlating with improvement in NSS 107.
Cell therapies were also investigated. Mesenchymal stem cell (MSC) transplantation was shown to reduce infarct volume 108 and improve NSS 109 in a TGFβ-dependent manner, and to increase serum TGF-β1 levels 110 when administered IV. Dental pulp stem cell injection also upregulated the TGF-β1 pathway, which correlated with reduced infarct size and improved NSS 111. Inversely, one study showed a downregulation of TGF-β1 following MSC injection (IV), though the effect of this on functional outcomes was not shown 112.
A wide range of other interventions were studied, with a majority showing a link between increased TGF-β1 expression and positive outcomes. Extracellular vesicles derived from oxygen and glucose deprived cells 113 upregulated the TGF-β1 pathway, improving NSS 113. Cerebral laser therapy (660 nm, 2.64 J/cm2) was shown to upregulate TGF-β1 in the serum, correlated with reduced nitric oxide (NO) synthase expression, suggested to reduce the generation of cytotoxic nitric oxide 114. TGF-β1 and the Smad signaling pathway were upregulated by urinary kallidinogenase 115, the neuroprotective agent N-butylphtalide 116, bovine myelin basic protein 117, tamoxifen 118, vitamin D3 119, R,S-ketamine120, coicis semen 21, nicotine 121 and β-1, 3-galactosyltransferase 2 122, all improving outcomes including infarct size and neurological function. Electroacupuncture was shown to increase TGF-β1 expression following tMCAO, correlated with increased expression of the apoptosis suppressor BCL-2 123. However, the opposite was shown in two studies; a reduction in TGF-β1 expression or signaling by pre-treatment with efonidipine 124 and by two Chinese herbal medicines 125 was correlated with improved outcome scoring.
Table 1 to be placed here – please find in Section 8: Tables larger than A4
4.6. TGF-β1 in pre-clinical hemorrhagic stroke studies
Three hemorrhagic stroke studies were performed without intervention. Of these, two suggested involvement of TGF-β1 in hemorrhagic stroke; showing that TGF-β1 was increased in a model of SAH 126 and that TGF-β1 expression correlated with the development of post-hemorrhagic hydrocephalus following IVH 127. In contrast, one study suggested that TGF-β1 expression was not induced in a blood injection model of ICH 128.
Nine hemorrhagic stroke studies (43%) modulated TGF-β1 directly. Studies directly applying TGF-β1 antagonists or inhibitors (6 studies, using anti-TGF-β1 antibodies, decorin, LSKL, SD208 and SB431542) reported significantly improved NSS and reduced ventricular enlargement 59–61,129,130 in models of SAH, ICH and GMH. These studies suggested a link between inhibiting TGF-β1 and improved outcome post-hemorrhagic stroke, typically by preventing fibrosis, especially in SAH models.
However, the role of TGF-β1 antagonism with oral anti-TGF-β1 drugs pirfenidone and losartan was unclear, with no effect noted in an IVH model 131. Exogenous TGF-β1 (3 studies) was only given as an intervention in ICH models and showed some protective effects, with reduced NSS and BBB leakage reported following injection of TGF-β1 to the striatum 132 and peritoneum 133. However, one study reported the induction of hydrocephalus following intrathecal injection of 6 ng TGF-β1, or TGF-β1-containing serum 134.
The anti-inflammatory agent minocycline was shown to exert a protective effect in an ICH model, with upregulation of TGF-β1 correlated with reduced NSS and cerebral oedema 85. Similarly, the cannabinoid receptor 2 (CBR2) agonist JWH-133 generated increased TGF-β1 expression and reduced NSS and brain water content in a SAH model 135. Interestingly, the opposite effect was noted in an IVH model, with JWH-133 being shown to down-regulate TGF-β1 and improve functional outcomes while reducing subarachnoid fibrosis and lateral ventricle size 62.
Simvastatin, a HMG-CoA reductase inhibitor, also exerted a protective effect, with reduced NSS and higher expression of TGF-β1 in the brainstem 136. Unrestricted somatic stem cell ICV infusion was also investigated, exerting a protective effect by increasing TGF-β1 protein in the CSF and RNA in the brain, correlated with increased cell proliferation 63.
Inversely, TGF-β1 expression correlated with negative outcomes in studies of the antioxidant resveratrol 137, the insulin sensitizer adiponectin 138 and an aryl hydrocarbon receptor agonist 139. Colchicine 140 generated no significant effect on TGF-β1 expression or relevant outcomes.
Table 2 to be placed here – please find in Section 8: Tables larger than A4
4.7. Risk of bias in pre-clinical stroke studies
The studies included in the present review typically showed low compliance with the SYRCLE risk of bias (RoB) tool for pre-clinical animal studies, with a low score indicating high risk of bias. The mean number of SYRCLE RoB criteria met was 3.90 across 89 studies, with two studies agreeing with zero of the criteria, and no studies complying with all 10 criteria (Figure 5). Complete data showing the total RoB score for each study included in the present work can be found in Additional File 1, Table 3.
{kind=link}