Result 1: Characterization of circDDX17 existence, subcellular distribution, and expression in GC cells and tissues
Utilizing the GEO database (https://www.ncbi.nlm.nih.gov/gds/), we identified differentially expressed circRNAs. CircRNA microarrays from GSE181769, GSE121445, and GSE100170 datasets encompass paired GC tissues and adjacent non-tumor tissues, along with the GSE202538 dataset containing GC cell lines and GES-1 cells. Overlapping the four circRNA microarrays facilitated the isolation of hsa_circ_0063331 (circDDX17) (Fig. 1A). Notably, both GC cells and tissues exhibited low expression levels of circDDX17. The transcript ddx17, which is cyclized from exons 2–5, has a length of 451 bp and is located on chromosome 22 (Fig. 1B). Amplification of circular transcripts employed divergent primers, while linear transcripts were analyzed using convergent primers. Moreover, circDDX17 exhibited resistance to RNase R treatment, confirmed by the inability of divergent primers to amplify circDDX17 from the cDNA of HGC-27 cells (Fig. 1C, D). Subsequent nuclear mass separation assay (Figs. 1E) and FISH analysis (Figs. 1F) demonstrated the cytoplasmic presence of circDDX17 in HGC-27 and MKN-45 cells.
Expression analysis revealed lower circDDX17 levels in gastric tumor tissues compared to 51 pairs of neighboring non-tumor tissues and normal tissues (Fig. 1G). Importantly, the expression of circDDX17 in tumor tissues was negative and substantially correlated with the size of tumor tissues and lymph node metastases in patients, as indicated by the clinicopathological analysis of circDDX17 (Table 1). Furthermore, circDDX17 gene expression in GC cell lines was notably diminished compared to GES-1 cells (Fig. 1H).
Table 1
The clinicopathological features of GC patients with differential expressions of circDDX17
| Clinical Parameters | Cases(51) | Expression of circDDX17 | P value |
| High (n = 14) | Low (n = 37) |
| Gender | | | | ns |
| Male | 38 | 10 | 28 | |
| Female | 13 | 4 | 9 | |
| Age (years) | | | | ns |
| > 65 | 37 | 12 | 25 | |
| ≤ 65 | 14 | 2 | 12 | |
| Tumor size (cm) | | | | < 0.05 |
| > 4 | 24 | 10 | 14 | |
| ≤ 4 | 27 | 4 | 23 | |
| Lymphatic Metastasis | | | | < 0.05 |
| Yes | 22 | 9 | 13 | |
| No | 29 | 5 | 24 | |
| TNM stage | | | | ns |
| I + II | 23 | 4 | 19 | |
| III + IV | 28 | 10 | 18 | |
Result 2: CircDDX17 promotes cell apoptosis, and inhibits cell proliferation, migration, and EMT GC cells
To elucidate the function of circDDX17 in GC, circDDX17 was either silenced in MGC-803 cells or overexpressed in HGC-27/MKN-45 cells. Successful modulation of circDDX17 expression was confirmed by qRT-PCR (Fig. 2A), with no alternation observed in ddx17 mRNA levels (Fig. 2B). Colony formation (Figs. 2C and 2D), CCK8 (Figs. 2E-2G), and flow cytometry (Fig. 2H) assays revealed that GC cells overexpressing circDDX17 decreased cell proliferation and promoted apoptosis, whereas silencing circDDX17 increased cell proliferation and inhibited cell apoptosis.
Investigating the role of circDDX17 in GC cell migration and EMT revealed that overexpression of circDDX17 significantly attenuated migration in HGC-27 and MKN-45 cells (Fig. 2I) while enhancing migration in MGC-803 cells (Fig. 2J). Transwell assays corroborated these findings, showing reduced migration in GC cells overexpressing circDDX17 (Fig. 2K) and enhanced migration upon circDDX17 silencing (Fig. 2L).
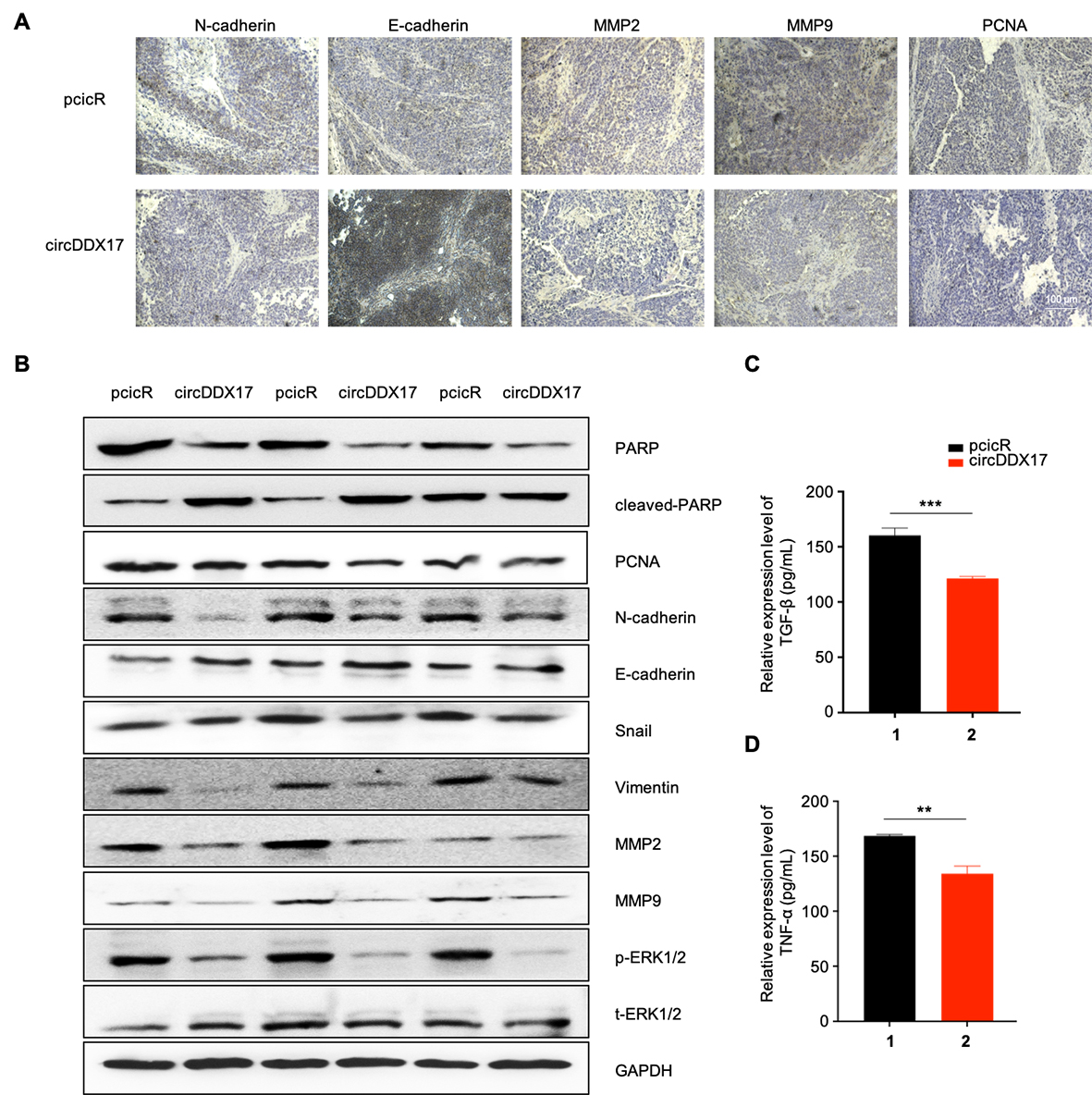
Western blot analysis of proliferation and apoptosis-related proteins (Fig. 2M) indicated that HGC-27 and MKN-45 cells overexpressing circDDX17 upregulated the expression of cleaved-PARP and BAX while suppressing the expression of BCL and PCNA. Conversely, circDDX17 silence in MDC-803 cells downregulated cleaved PARP and BAX and reduced BCL and PCNA expression. Similarly, Western blot data showed that circDDX17 decreased EMT, with the opposite effect observed upon circDDX17 silencing (Fig. 2N). In vivo, circDDX17-overexpressing reduced GC cell proliferation, migration and EMT, by IHC and Western blot, and decreased serum levels of TGF-β and TNF-α expression by ELISA (Figure S1).
Results 3 CircDDX17 functioned as a miR-1208 and miR-1279 sponge
Our investigation delved into whether circDDX17 modulated GC cell behavior by acting as a sponge for specific miRNAs, a pivotal role attributed to circRNAs. Initially, two miRNAs, miR-1208 and miR-1279, were identified from circBank (http://www.circbank.cn) using bioinformatics analysis, both potentially interacting with circDDX17. qRT-PCR analysis unveiled a negative regulatory effect of circDDX17 on miR-1208 and miR-1279 expression in GC cells (Figs. 3A and 3B). The full-length circDDX17-wild-type (wt) and circDDX17-mutant (mut) sequences were then cloned into the luciferase vector in the absence of miR-1208 or miR-1279 binding sites (Fig. 3C). Luciferase assays confirmed that miR-1208 and miR-1279 significantly decreased the luciferase activity of circDDX17-wt but not circDDX17-mut compared to the miR-NC group (Figs. 3D and 3E).
Further exploration of miR-1208 and miR-1279 expression in GC cell lines, adjacent non-tumor tissues, and gastric tumor tissues was performed using qRT-PCR. Results revealed elevated expression levels of miR-1208 and miR-1279 in GC cell lines and gastric tumor tissues compared to surrounding non-tumor tissues (Fig. 3F–3I). Table S1 shows a correlation between miR-1208 expression level and lymph node metastasis, and Table S2 shows a correlation between miR-1279 expression level and tumor size.
Results 4 miR-1208 and miR-1279 reversed the ability of circDDX17 on GC cell apoptosis, proliferation, migration, and EMT
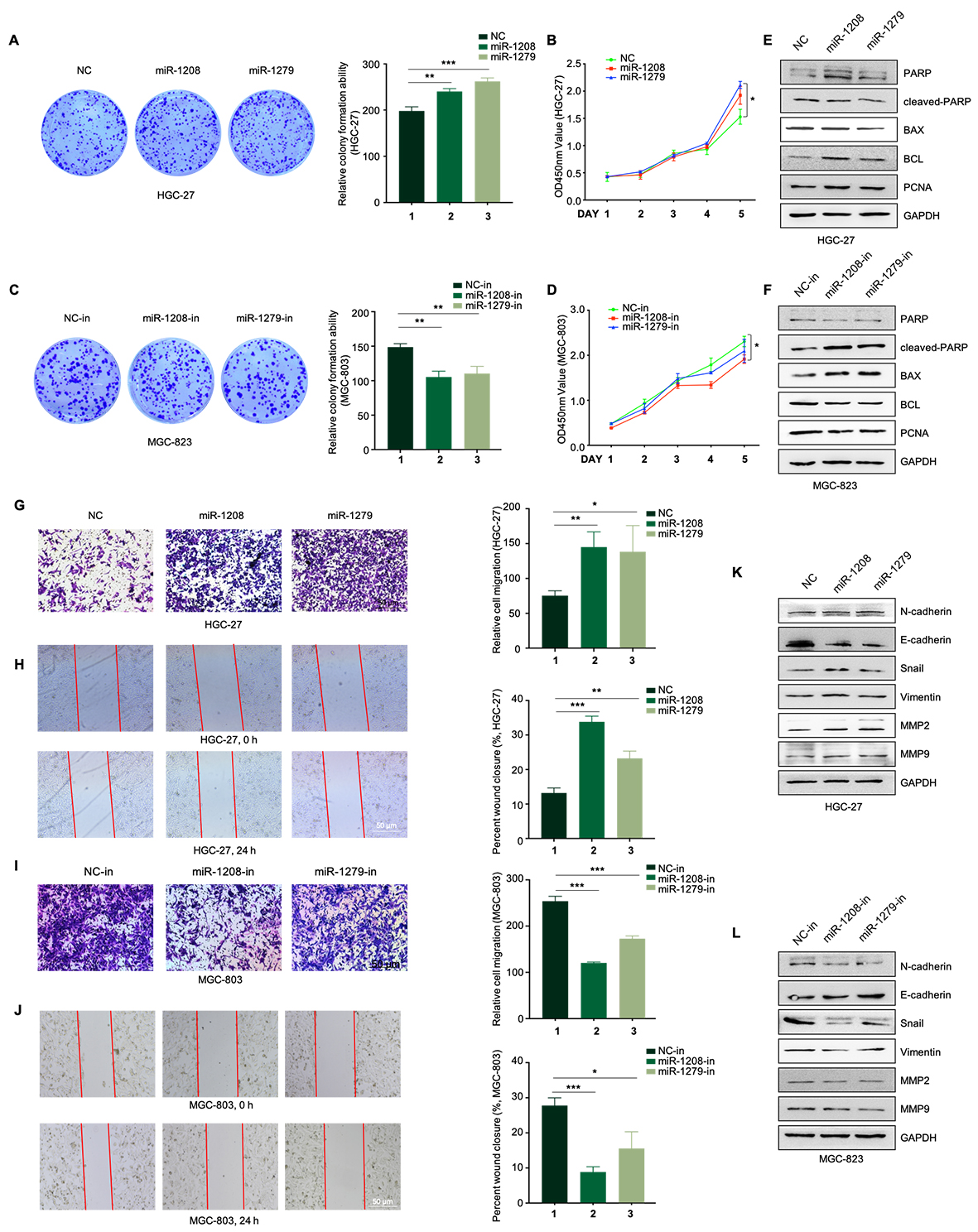
miRNAs play pivotal roles in GC pathogenesis, prompting an investigation into their functions through either overexpressing or silencing in GC cells. Colony formation, CCK8, Transwell, wound healing, and Western blot collectively revealed that miR-1208 and miR-1279 mimics enhanced GC cell proliferation, migration, and EMT, while miR-1208 and miR-1279 inhibitors suppressed GC cell proliferation (Figure S2), affirming their tumor-promoting impact on GC.
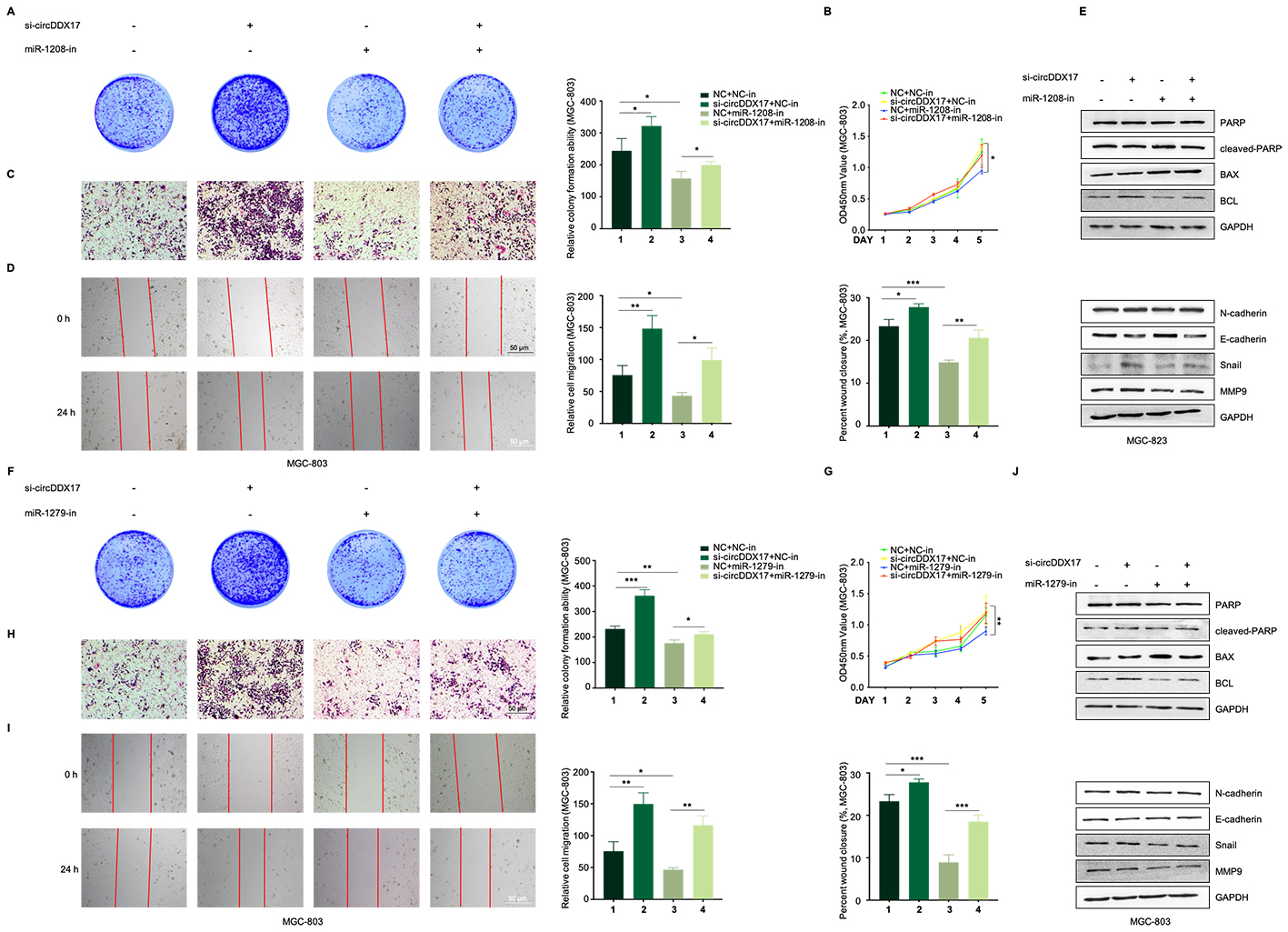
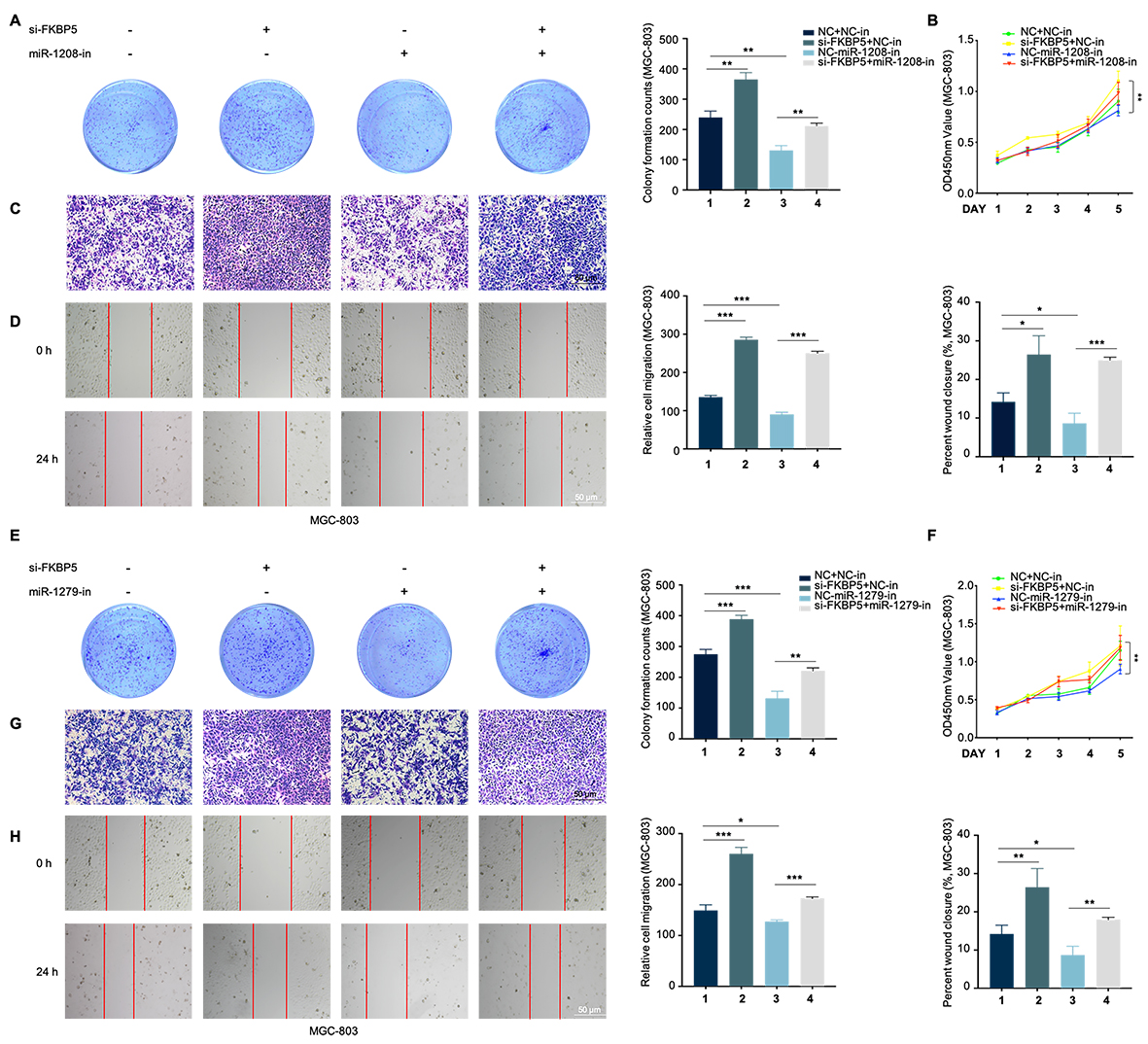
To ascertain if circDDX17 exerted biological functions by sequestering miR-1208 and miR-1279, rescue experiments were conducted wherein circDDX17 and miR-1208/miR-1279 mimics co-transfected in HGC-27 cells. Colony formation (Fig. 4A and 4F), CCK8 assay (Fig. 4B and 4G), and Western blot assays demonstrated that miR-1208/miR-1279 reversed the trend of circDDX17 in apoptosis and proliferation (Fig. 4C and 4H). Moreover, Transwell, wound healing, and Western blot data indicated that miR-1208/miR-1279 reversed the trend of circDDX17 on migration and EMT (Fig. 4D-4E and 4I-4J). Additionally, si-circDDX17 enhanced proliferation and migration in MGC-803 cells, while miR-1208-in or miR-1279-in reduced proliferation and migration ability (Figure S3).
Results 5. CircDDX17 regulates FKBP5 by miR-1208 and miR-1279
To elucidate the downstream mechanisms of miR-1208 and miR-1279 downstream factors, bioinformatics tools (Targetscan and miRDB) were employed to predict the potential targets of miR-1208 and miR-1279. As shown in Fig. 5A, FKBP5 acts as the miR-1208 and miR-1279 target gene. Expression analysis in HGC-27 and MGC-803 cells revealed that miR-1208 and miR-1279 negatively regulated FKBP mRNA (Fig. 5B and 5C) and protein (Fig. 5D) expression. The expression of FKBP5 mRNA in human tissues was also identified. Compared to control tissues, the expression of FKBP5 was significantly downregulated in tumor tissues (Fig. 5E). Moreover, the expression of FKBP5 in tumor tissues was negatively and significantly associated with the size of tumor tissues (Table 2). IHC data confirmed low FKBP5 expression in gastric tumor tissues (Fig. 5F). Additionally, the expression of protein FKBP5 was lower in the GC cell line than in GES-1 cells (Fig. 5G), suggesting FKBP5 is a target gene of miR-1208 and miR-1279.
Table 2
The clinicopathological features of GC patients with differential expressions of FKBP5
| Clinical Parameters | Cases(51) | Expression of FKBP5 | P value |
| High (n = 15) | Low (n = 36) |
| Gender | | | | ns |
| Male | 38 | 10 | 28 | |
| Female | 13 | 5 | 8 | |
| Age (years) | | | | ns |
| > 65 | 37 | 9 | 28 | |
| ≤ 65 | 14 | 6 | 8 | |
| Tumor size (cm) | | | | < 0.05 |
| > 4 | 24 | 11 | 13 | |
| ≤ 4 | 27 | 4 | 23 | |
| Lymphatic Metastasis | | | | ns |
| Yes | 22 | 8 | 14 | |
| No | 29 | 7 | 22 | |
| TNM stage | | | | ns |
| I + II | 23 | 5 | 18 | |
| III + IV | 28 | 10 | 18 | |
Table S3
| Primers | Sequences(5'-3') |
| CircDDX17-F | ATTTCCGTTGGCTCTTAGTG |
| CircDDX17-R | CCTCTTGCTCCAAATGATTG |
| circDDX17-mRNA-F | ATTTGGAGCAAGAGGTGGTG |
| circDDX17-mRNA-R | CACTAAGAGCCAACGGAAAT |
| GAPDH-F | AGGTGAAGGTCGGAGTCAAC |
| GAPDH-R | GGGTGGAATCATATTGGAACA |
| GAPDH-circ-F | TTGCCCTCAACGACCACTTT |
| GAPDH-circ-R | ACCAAATCCGTTGACTCCGA |
| U6-F | CTCGCTTCGGCAGCACA |
| U6-R | AACGCTTCACGAATTTGCGT |
| 18s-F | ACACGGACAGGATTGACAGA |
| 18s-R | GGACATCTAAGGGCATCACA |
| miR-1208-F | CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGTCCGCCTG-3 |
| miR-1208-R | ACACTCCAGCTGGGTCACTGTTCAGACA |
| miR-1279-F | CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGAGAAAGAA |
| miR-1279-R | ACACTCCAGCTGGGTCATATTGCTT |
| FKBP5-F | AACCCCCAGAATAAGGCTGC |
| FKBP5-R | TTGGCGTATATCCTGCGGTC |
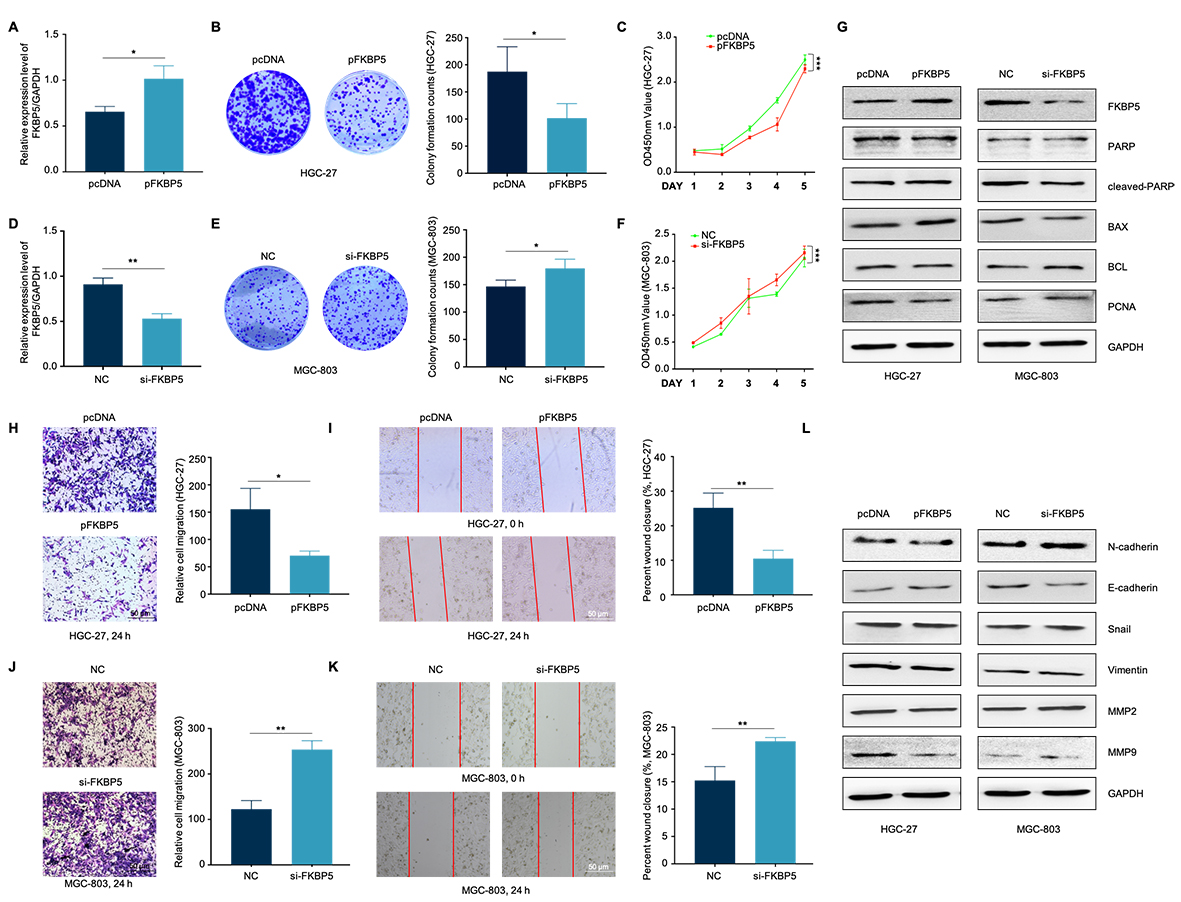
Further investigation into the impact of miR-1208/miR-1279-FKBP5 on GC cell phenotype revealed that FKBP5 overexpression in HGC-27 cells reduced the healing ability and migratory capacity, and to some degree, decreased proliferation and formed smaller clones compared to control cells. Contrary to the above results, si-FKBP5 promotes GC cell migration and proliferation (Figure S4).
Rescue experiments were conducted to analyze how FKBP5 influenced the effects of miR-1208/miR-1279 on GC development. Functionally, FKBP5 abolished the effects of miR-1208/miR-1279 on GC cell proliferation, migration, and apoptosis (Fig. 5H-5J, 5N-5P). Furthermore, the abnormal FKBP5 levels within GC cells restored miR-1208/miR-1279 role in migration and EMT (Fig. 5K-5M, 5Q-5S). Conversely, miR-1208-inhibitor/miR-1279-inhibitor reversed the effects of si-FKBP5 on GC (Figure S5).
Results 6. FKBP5 regulated GC progression by the ERK1/2 pathway
FKBP5 was initially identified in the HeLa cell cDNA library13. According to the Oncomine database, FKBP5 expression varies across different cancer types. For example, FKBP5 is highly expressed in brain cancer, prostate cancer, and lymphoma but is expressed at low levels in pancreatic14, colon, testicular, and gastric cancers 15. Our study found that FKBP5 expression was low in both GC cell lines and tissues and is inversely correlated with tumor size in patients with GC. Overexpression of FKBP5 decreased GC cell migration, EMT, and proliferation while promoting apoptosis. Abnormal activation of the ERK pathway contributes to the growth of various tumor types and affects cellular functions like metastasis, cell survival, and proliferation 16.
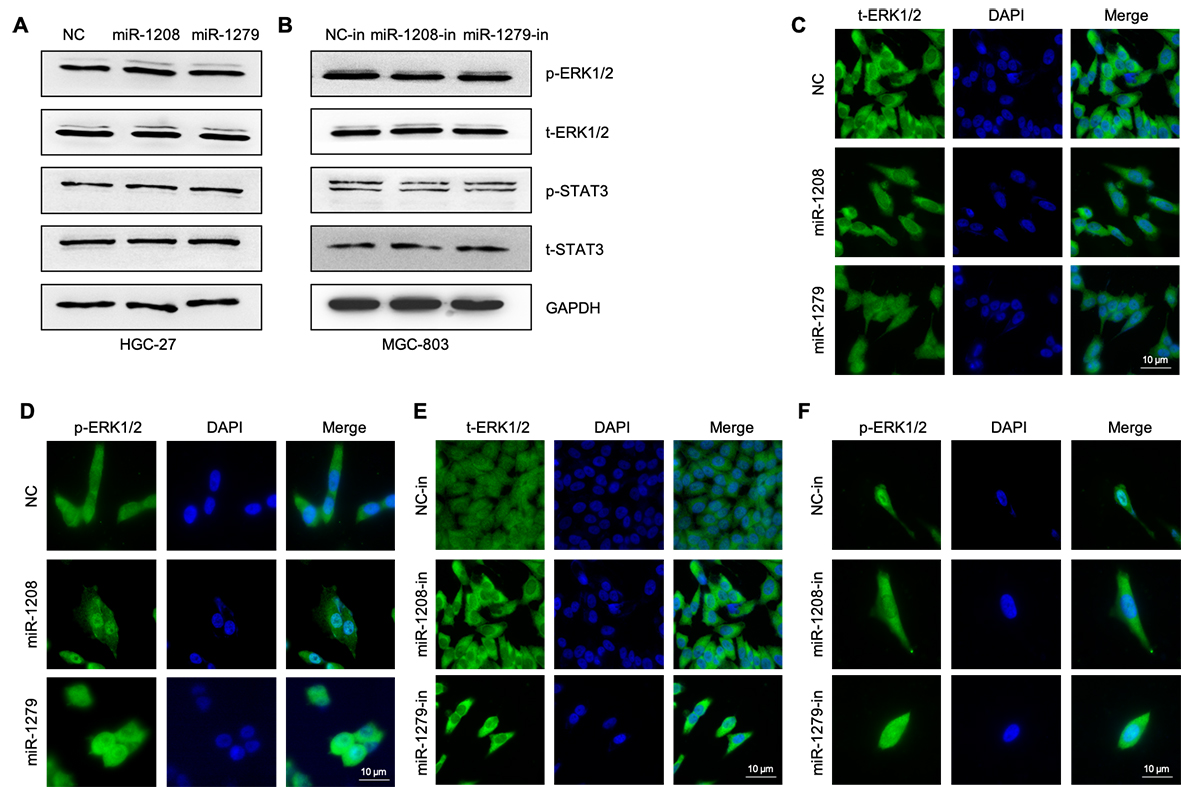
To explore the underlying mechanism by which FKBP5 facilitates GC progression, we used bioinformatics tools (STRING, https://cn.string-db.org/cgi/network) to predict interacting proteins and identified ERK1/2 as a target protein of FKBP5. To confirm the interaction between ERK1/2 and FKBP5, we performed IP/Western blot analysis using an anti-FKBP5 antibody. Results showed that exogenous FKBP5 and ERK1/2 bind in HGC-27 and MKN-45 cells (Fig. 6A). An IF assay and confocal imaging revealed that FKBP5 and ERK1/2 co-localized in the cytoplasm of HGC-27 cells (Fig. 6B). When ERK1/2 was inactivated in HGC-27 cells using PD98059, Western blot (Fig. 6C) and IF (Fig. 6D-6F) data revealed that FKBP5, t-ERK1/2, and p-ERK1/2 were significantly decreased. This suggests that PD98059 can concurrently reduce the expression of FKBP5 and ERK1/2. Figures 6G illustrate that FKBP5 overexpression decreased the expression of p-ERK1/2/t-ERK1/2 and p-STAT3/t-STAT3 in intracellular. Figures 6H and 6I show that the overexpression of FKBP5 inhibited the expression of p-ERK1/2 in cells, with the inhibitory effect being particularly noticeable in the nucleus. Conversely, MGC-803 knockdown FKBP5 improved p-ERK1/2 activated in the nucleus and elevated p-ERK1/2/t-ERK1/2 and p-STAT3/t-STAT3 (Fig. 6J-6L).
To further analyze how circDDX17 affects the ERK1/2 and STAT3 pathway via the miR-1208/miR-1279/FKBP5 axis, we examined the effects of circDDX17 overexpression or knockdown in GC cells. circDDX17 overexpression decreased ERK1/2 and STAT3 phosphorylation in HGC-27 and MKN-45 cells (Fig. 6M), whereas circDDX17 knockdown enhanced ERK1/2 and STAT3 phosphorylation in MGC-803 cells (Fig. 6N). When HGC-27 and control cells were overexpressed and injected into the nude mice, FKBP5 was upregulated in the tumor tissue (Fig. 6O); however, ERK1/2 phosphorylation were downregulated (Fig. 6P). Furthermore, IF results showed that circDDX17 knockdown boosted ERK1/2 and STAT3 activation in the nucleus (Fig. 6Q-6T).
Results 7. CircDDX17 encodes the 63aa tumor-suppressed protein
Based on our previous results, we established that circDDX17 suppresses proliferation, migration, and EMT in GC cells. We further explored whether circDDX17 exerts its function by encoding a peptide. circDDX17 was mainly located in the cytoplasm, indicating the potential for protein encoding. The ability and sequence of the polypeptide encoded by circDDX17 were predicted using the public database ORF Finder (www.ncbi.nlm.nih.gov/orffinder), which revealed that circDDX17 could encode a polypeptide, with a 192nt (63aa) segment identifies as a potential encoding sequence (Fig. 7A).
According to the online prediction results, the peptides encoded by circDDX17 are expressed in eukaryotic plasmids. A novel circDDX17 vector containing the GFP sequence was constructed to further assess the protein-coding ability of circDDX17. The circularization of circDDX17 creates a tandem start codon ‘AUG’, initiating translation with overlapping genetic codes. To differentiate the role of the circDDX17-63aa peptide from circDDX17, we constructed several GFP-tag vectors for circDDX17 (Fig. 7B). Transfection with the Lv- circDDX17 vector and Lv- circDDX17 -mut vector both successfully resulted in the overexpression of circDDX17 and the GFP-tagged proteins were detected using Western blot after extracting total cellular proteins from transfected HEK-293T cells after 48 h. (Fig. 7C). Lv-circDDX17 and Lv-circDDX17-63aa successfully expressed GFP-tag; however, the Lv-vector and Lv-circDDX17-mut did not express the GFP-tag. Vectors transfected in HGC-27 cells, GFP-tag expression on Lv-circDDX17 and Lv-circDDX17-63aa transfected cells, and GFP-tag were observed in the cytoplasm (Fig. 9D). SDS-PAGE and Western blot analysis identified the circDDX17-63aa protein at the 10-kDa band, confirming its translation from circDDX17 (Figs. 7E and 7F).
To further explore the biological function of circDDX17-63aa, we transfected HGC-27 cells within the four aforementioned vectors. Overexpression of wild-type circDDX17-63aa decreased proliferation ability, as evidenced by colony formation (Fig. 7G and 7H) and CCK8 assay (Fig. 7I). Moreover, circDDX17-63aa suppressed GC cell migration, evidenced by Transwell (Fig. 7J and 7L) and wound healing assay (Fig. 7K and 7M).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}