In this study, we re-evaluated the anatomical structure of the human GM, updating information on its insertion. The large superior portion of the GM was mostly inserted into the gluteal tuberosity via the thick plate-like tendon, and the small inferior portion was inserted into the complex including the thick plate-like tendon, LFIS, and proximal tendon of the vastus lateralis. The GM superior portion can generate a large muscle force because of its significantly larger PCSA. Conversely, the inferior portion has a significantly smaller PCSA, although it had good muscle contractility, as evidenced by its significantly longer muscle fiber length, implying that more sarcomeres are arranged per muscle fiber.
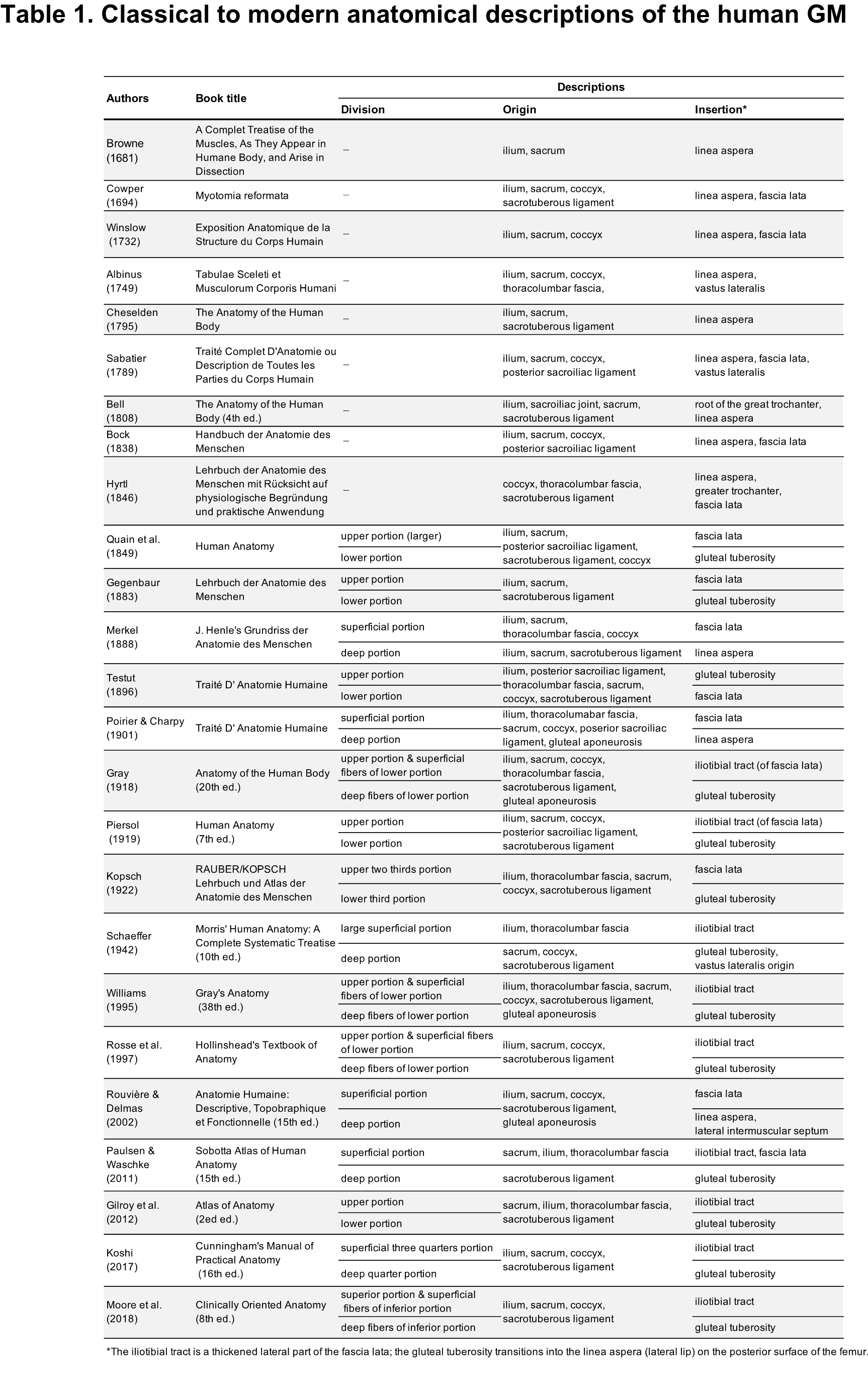
Historically, American, British, and German literature have distinguished the GM into superior (large) and inferior portions based on structural differences in the distal region, whether the fascicles were inserted into the iliotibial tract (or the fascia lata) or the femur, since the 19th century1–3,21−39 (Table 1). From around 1900, there was a tendency to distinguish GM compartments as superficial (inserting into the iliotibial tract) and deep (inserting into the femur). Current anatomical descriptions incorporate these criteria for distinguishing GM portions. However, inconsistency exists: the deep fibers of the inferior portion are typically described as attaching to the femur, this contrasts with both some existing descriptions and our findings. In French literature, Testut (1896) described the GM insertion similarly to our findings, noting the inferior fascicles attach mostly to the fascia lata, with the remaining fascicles terminating on the gluteal tuberosity of the femur whereas it was not shown in any Fig. 40. Poirier and Charpy (1901) illustrated the actual structure of the distal GM tendon with accurate drawings unique to previous reports, although this tendon was recognized as terminating at the fascia lata41. In recent French anatomy texts, such as Rouvière and Delmas (2002), descriptions not found in anatomy texts in English- or German-speaking countries have been eliminated42. The reason for this misunderstanding regarding the insertion of the human GM is unclear, although partial adherence of the superior portion to the iliotibial tract is a major contributing factor. Additionally, from all the descriptions and figures in each previous report, it is understandable that GM distal end is not pursued adequately in in situ observation, which may create long-standing misconceptions regarding GM insertion and division. The present structural re-evaluation based on in situ and isolated muscle specimen analyses enabled us to structurally characterize the distal region of the human GM and to establish the division of the GM into large superior and small inferior portions.
From a developmental anatomy perspective, Tický and Grim (1985) reported that the human GM comprised two fused fetal muscles: the fetal GM proper (superior) and the fetal coccygeofemoralis (inferior), which are constantly observed in human embryos and fetuses43. Shiraishi et al. (2018) also found that the GM divides into superior and inferior portions in the early fetal period, with the inferior portion developing approximately 1 week later than the superior portion, and the two portions unite to form an adult form44. If the superior and inferior portions of the fetal GM correspond to those of the adult GM, the division of the adult GM (and its structural differences) is presumably attributable to the developmental pattern in which the GM arises from two different muscle primordia.
Regarding functional anatomy, the GM is known for extending the hip joint and contributing to other hip joint motions, such as hip abduction and external rotation6–8. Previous anatomical and kinesiological literature have described that the GM contributes to the movement and stabilization of the knee joint via the iliotibial tract1–3,5,45. However, our findings suggest that any GM contribution to knee joint movement is minimal. A recent kinesiological study using electromyography and ultrasound tomography also showed that GM contraction does not contribute to the movement/deviation of the iliotibial tract14, supporting our present findings.
We found that the superior portion, with a large PCSA, was inserted into the femur with a strong tendon. The inferior portion was speculated to indirectly assist the superior portion by tensing the distal tendon of the superior portion. Additionally, the small accessory fibers of the superior portion that united the iliotibial tract are presumably unworkable for tibial movement because the iliotibial tract is fixed to the shaft of the femur by the LFIS, as previously described by several authors38,46,47. Our findings indicate that GM contributes to hip joint movement. This aligns with kinetic studies by Preece et al. (2008) who found an unclear link between GM activity and tibial rotation during gait48, and Besomi et al. (2022) who reported no correlation between GM activity and iliotibial tract displacement14. These findings support anatomical observations and functional suggestions in terms of kinesiology. Conversely, biomechanical studies of lower limb movement using muscle modeling, which are presented based on previous GM descriptions, reported that GM greatly contributes to the movements of both the hip and knee joints as a powerful extensor49,50. Based on our precise anatomical data, kinesiological data on the GM should be re-interpreted. This will significantly improve our understanding of human GM function.
{kind=link}