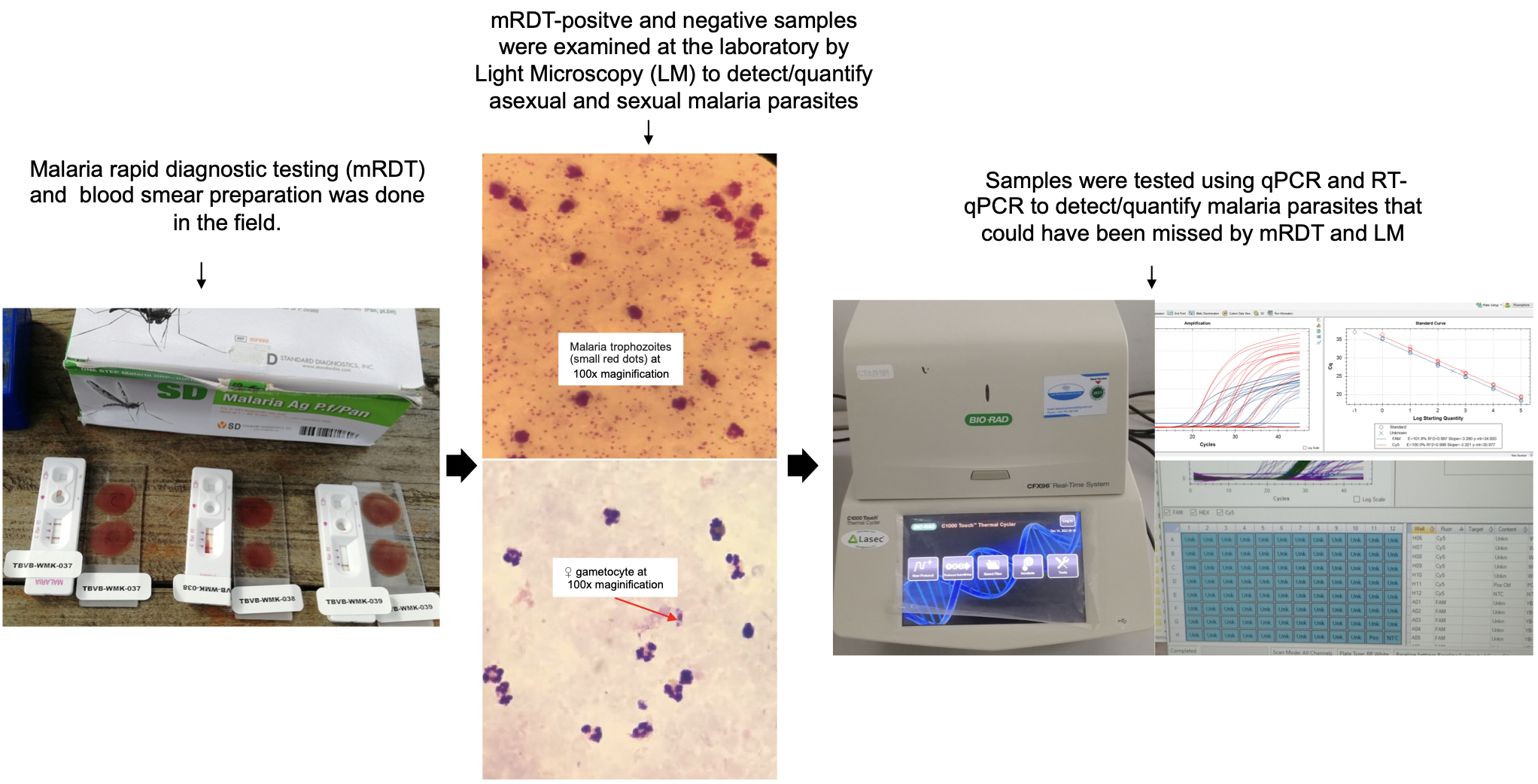
Better understanding of the infectious reservoir of Plasmodium falciparum better informs the development and evaluation of new tools that can interrupt malaria transmission. Here, we determined malaria parasite prevalence and density in asymptomatic school-age children, adolescents and adults from Bagamoyo district in Tanzania. In line with common practices in many African countries, we utilized malaria rapid diagnostic tests (mRDTs) and microscopy to detect malaria infections in the study. Additionally, we employed sensitive qPCR assays to ensure the detection of asymptomatic infections that may be missed by mRDTs and microscopy. Our findings showed that both malaria positivity and gametocyte prevalence were significantly higher in children and adolescents compared to adults.
We observed that microscopy missed more than one-third (33.6%) of infections, whereas mRDT missed nearly 40% of the qPCR-positive infections. These findings corroborate previous studies in Tanzania [31], Nigeria [35], and Ghana [36], where significant proportions of qPCR-positive infections were missed by microscopy and mRDT. The missed infections recorded by microscopy and mRDT in the current study could be attributed to submicroscopic parasites, which tend to persist for several months without any symptoms. Furthermore, the observed mRDT false negatives may have been as result of HRP-2 gene deletions, which is prevalent in Tanzania [37]. The missed infections are of great concern since failure to identify malaria-carriers may lead to continued transmission and increase in the malaria burden, particularly in low-transmission areas. The observed false positives by mRDT may be as a result of persistence of HRP2 antigen after successful antimalarial treatment. The persistent HRP2 positivity is more common when treatment is done with artemisinin combination therapy and may last for 2–20 days post-treatment [38].
There have been previous reports of relatively high gametocyte prevalence in school-age children in Bagamoyo district than what we observed; one study in Kiwangwa village reported a gametocyte prevalence of 14% [39], and another study in Buma and Yombo reported a gametocyte prevalence of 18% [23].Given the difference in study villages, a direct comparison between studies may not be appropriate. Nonetheless, the difference between the gametocyte prevalence in our study and previous reports could be reflective of heterogeneity in malaria transmission and carriage rates in the population. Futhermore, a large proportion of gametocyte carriers in our study were adults, indicating a shift in the age-distribution of gametocyte carriage. This shift could be attributed to intensive efforts to reduce and prevent malaria in children than in adults in recent years [40, 41]. As a tradition, men and young boys in our study area are more engaged in malaria-risk activities such as; fishing at night, animal grazing among others. This could explain why a higher gametocyte prevalence was obseverd in male participants compared with their female counterparts.
About 24% malaria-positive participants in our study were found to harbor P. falciparum gametocytes, underscoring the prevalence of these sexual-stage parasites in a predominantly asymptomatic population where malaria infections are often submicroscopic [23, 28, 39]. These results align with findings from several studies conducted in other malaria-endemic areas [42–45], which have reported similar age distributions. The overall gametocyte sex ratio in the current study is similar to what has been observed in other endemic areas [12]. The gametocyte sex ratio in adult gametocyte carriers of our study is an indication that adults could also be playing a significant role in malaria transmission. The gametocyte sex ratio, which is; the proportion of male gametocytes, is a useful predictor of the malaria infectiousness of human host and their potential to transmit malaria parasites [46]. Although the likelihood of mosquito infection largely depends on gametocyte density, parasite fertilization in the mosquito requires sufficient numbers of male and female gametocytes to be present in the mosquito blood meal [47]. Quantifying both male and female gametocytes in human hosts allows a better prediction of infectiousness than meaurement of the total gametocytes or the more abundant female gametocytes [12].
One third of identified gametocyte carriers were submicroscopic, highlighting the need for molecular testing in malaria surveillance and evaluation of interventions [28, 42]. A recent study in Bagamoyo correlated gametocyte presence with infectiousness to mosquitoes, indicating that 95% of all oocyst-infected mosquitoes after direct membrane feeding assays were due to submicroscopic malaria infections in school-age children [23]. While further prospective studies are warranted to fully elucidate the age-specific contribution to mosquito infection, particularly given the similar gametocyte density observed across age groups, our current findings hold important implications for community-level malaria control efforts. Notably, adults, like school-age children, constitute a substantial proportion of gametocyte carriers in the study area, suggesting that interventions focused solely on children may not suffice to effectively interrupt malaria transmission.
{kind=link}