Early detection and intervention play crucial roles in enhancing the clinical outcomes of MM patients [29, 30], emphasizing the pressing need to identify potential non-invasive biomarkers. Considering the intra-tumoral heterogeneity and systemic changes induced by hematological malignancy, the analysis of blood samples offers insights into the overall phenotype of MM. Moreover, in comparison to invasive detection methods such as bone marrow puncture or biopsy, an ideal biomarker for MM clinical diagnosis should exhibit optimal sensitivity and specificity when obtained from patients through minimally invasive means, such as blood.
Prior studies have explored metabolic biomarkers of MM in blood [31–33], indicating that molecular predictive classifiers could offer valuable insights for future targeted MM therapy. However, these studies focused primarily on identifying and describing the metabolic landscape of MM on the basis of single or specific clinical factors. In this retrospective metabolomics analysis of MM, we sought to address three key questions: (i) which patients could be screened early, (ii) which patients could be precisely staged, and (iii) which patients could benefit from chemotherapies. Notably, we observed a consistent decline in lactic acid and leucine levels in the plasma of MM patients at diagnosis, staging, and prognosis, suggesting that these metabolites are potential plasma biomarkers associated with active MM disease.
To the best of our knowledge, this is the most comprehensive analysis demonstrating the extent of metabolic reprogramming in MM. Importantly, the samples included in this study are real-world samples without strict enrollment criteria, demonstrating the robustness of our analysis. Following univariate analyses, we identified a biomarker panel that includes lactic acid and leucine.
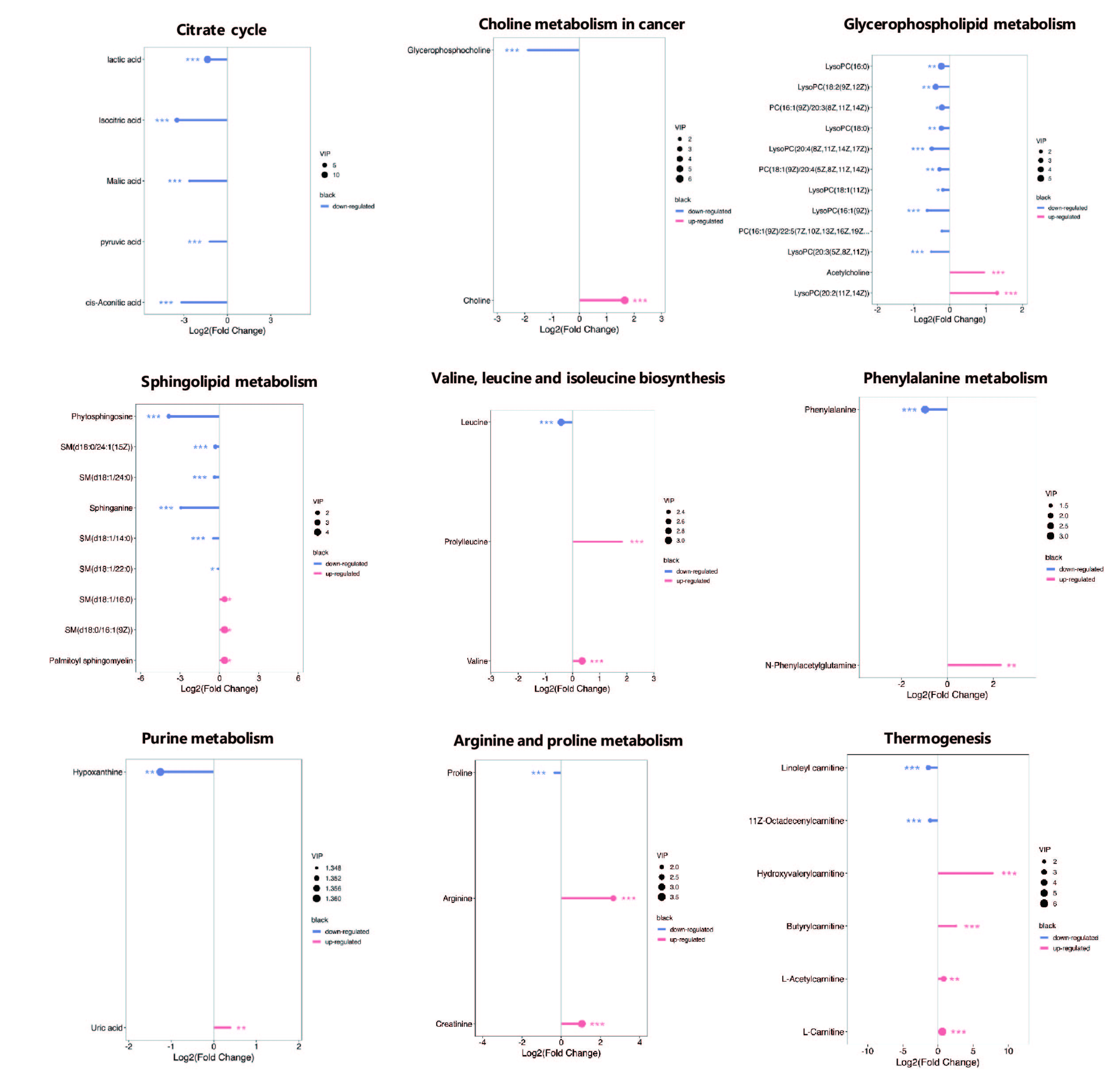
Lactic acid is often considered a marker of the “Warburg effect” in tumor cells [34, 35]. Warburg's observation revealed that, unlike most normal cells, tumor cells tend to ferment glucose to lactate even in the presence of sufficient oxygen to support mitochondrial oxidative phosphorylation. The acidic microenvironment formed by lactic acid is conducive to the rapid growth and distant metastasis of tumor cells. Wiled et al. [35] reported that lactate is also supplied to cancer cells from the surrounding environment, referring to this phenomenon as the reverse Warburg effect. Our observation of a decline in lactate levels in MM patients suggests that the reverse Warburg effect might be applicable to the microenvironment in MM, as previously reported [36]. Furthermore, in patients with MM who achieve complete remission, the increase in lactate concentration is particularly pronounced [22]. Notably, the elevated level of lactate dehydrogenase (LDH), the enzyme catalyzing the conversion of pyruvate to lactate, serves as a marker of poor prognosis at the time of MM diagnosis. An increase in LDH is associated with worse overall survival (OS), progression-free survival (PFS), aggressive disease, and a higher tumor burden [35, 36]. Correspondingly, our research demonstrates that a decreased lactate level is closely related to MM initiation and progression, and it is expected to become an important biomarker for clinical diagnosis and treatment of MM in the future.
Another biomarker, leucine, one of the branched chain amino acids (BCAAs), is an important amino acid that plays crucial roles in the body. Compared with the HC group, MM patients presented lower levels of leucine. In another related study, the amino acid profiles of MM presented relatively low concentrations of leucine [33]. Furthermore, the concentration of essential amino acids, especially leucine, was significantly decreased in MM patients [39], which was consistent with our findings. Therefore, leucine appears to be a potential biomarker that should be evaluated in future studies addressing the diagnosis, staging, follow-up, prognosis, and treatment of MM.
Interestingly, a recent study identified genes related to lactic acid and BCAAs metabolism as potential prognostic biomarkers independently associated with the overall survival of MM patients [41], which strongly supports our conclusion. Overall, we hypothesize that the diagnostic value may be improved by combining the examination of clinical indicators, lactate, and leucine levels.
Metabolic pathways constitute a highly organized network of sequential chemical reactions in an organism, playing a vital role in maintaining the energy and material balance necessary for life processes. Deeper insights into how these abnormalities disrupt normal metabolic pathways will aid in better prevention, diagnosis, and treatment of associated diseases. Gonsalves et al. [33] reported that BCAAs metabolism, tryptophan metabolism, phospholipid metabolism, and nucleotide turnover were potentially affected by MM, whereas Chanukuppa et al. [24] reported alterations in pyrimidine metabolism, purine metabolism, amino acid metabolism, nitrogen metabolism, sulfur metabolism, and the citrate cycle. Wei et al. reported significant metabolic disorders in 46 pairs of pre- and post-therapy MM patients, specifically in arginine, proline and glycerophospholipid pathways [26]. These findings confirmed the vital role of certain metabolites and metabolic pathways in the pathogenesis of MM.
The citric acid cycle (TCA cycle), also known as the Krebs cycle, is a crucial biochemical pathway that occurs in the mitochondria of eukaryotic cells. It generates energy in the form of ATP by coupling the breakdown of substrates to the phosphorylation of ADP. The TCA cycle is a highly regulated process that responds to changes in nutrient availability and energy demand [41]. It can be upregulated during periods of high energy demand or downregulated during periods of nutrient abundance or energy excess. We found that isocitric acid, malic acid, pyruvic acid, and cis-aconitic acid in the TCA cycle were significant downregulated in MM patients, possibly suggesting that the TCA cycle was inhibited. This finding is consistent with Warburg's suggestion that even when oxygen is sufficient, tumor cells rely on massive glucose uptake, converting it to lactate for energy [36].
Lipids serve not only as essential components of cell membranes and energy storage systems but also as crucial signaling molecules that regulate biological processes under both normal and diseased conditions. In our study, we observed significant disturbances in choline metabolism, glycerophospholipid metabolism, and sphingolipid metabolism in MM patients. Choline, an integral part of acetylcholine synthesis and a precursor to phospholipid synthesis [42], undergoes transformation into phosphocholine, which is coupled with diacylglycerol to form phosphatidylcholine (PCs), a major component of cell membranes. Therefore, choline is considered to reflect the intensity of cell membrane synthesis. Our findings revealed a significant increase in choline and acetylcholine in the MM group compared with those in the HC group, whereas the abundances of PC(16:1(9Z)/20:3(8Z,11Z,14Z), PC(18:1(9Z)/20:4(5Z, 8Z, 11Z, 14Z), and PC(16:1(9Z)/22:5(7Z, 10Z, 13Z, 16Z.19Z)) were notably decreased in the MM group compared with those in the HC group. High choline uptake and downregulated PCs are believed to lead to the hydrolysis necessary for forming lipid messengers, responsible for the replication of clonal plasma cells and tumor dissemination [42]. Members of the lysophosphatidylcholines (LPCs), including LysoPC(16:0), LysoPC(18:2(9Z, 12Z)), LysoPC(18:0), LysoPC(20:4(8Z,11Z,14Z,12Z)), LysoPC(18:1(11Z)), LysoPC(16:1(9Z)), LysoPC(20:3(5Z,8Z,11Z)), and LysoPC(20:2(11Z, 14Z)), exhibited decreased levels in MM patients, consistent with previous research [43]. LPCs are crucial LDL/bioactive lipids that contribute to the inflammatory impact of oxidized LDL on endothelial cells. They are involved in inflammatory stimuli, and promote the release of IL-6 and other inflammatory factors, ultimately contributing to the development and progression of MM [25, 45]. Sphingolipids (SPs), another family of bioactive lipids with a structural role in the plasma membrane, have products of their metabolism (sphingosine, sphingosine-1-phosphate, ceramides, ceramide-1-phosphate) that play crucial roles in MM migration and adhesion, survival and proliferation, as well as angiogenesis and invasion [44]. In our study, most SPs exhibited significantly decreased levels in plasma, suggesting that SPs' hydrolysis can be part of the systemic metabolic regulation/reprogramming of MM.
Amino acids play an essential role in the synthesis of various biomolecules necessary for cell proliferation [46]. Moreover, targeting amino acid metabolism has been proposed as a potential cancer therapy, highlighting the importance of amino acid metabolism in cancer [47, 48]. Alterations in plasma amino acid profiles are relatively common in MM processes [23–26, 42]. In the present study, the main perturbed amino acid metabolism pathways in MM plasma were valine, leucine, and isoleucine biosynthesis, phenylalanine metabolism, and arginine and proline metabolism.
Leucine, valine and isoleucine, classified as branched-chain amino acids (BCAAs), are crucial for human life and are particularly involved in stress, energy and muscle metabolism [48]. BCAAs follow different metabolic routes, with valine exclusively contributing to carbohydrates (glycogenic), leucine solely to fats (ketogenic) and isoleucine being both a glucogenic and a ketogenic amino acid. The catabolism of valine begins with the removal of the amino group by transamination, producing alpha-ketoisovalerate, an alpha-keto acid, which is converted to isobutyryl-CoA through oxidative decarboxylation by the branched-chain alpha-ketoacid dehydrogenase complex. This is further oxidized and rearranged to succinyl-CoA, which can enter the TCA cycle. The elevated level of valine in MM patients may be due to the inhibited TCA cycle, as previously demonstrated.
Furthermore, we observed significant increases in arginine and creatinine levels in the MM group compared with those in the HC group. Arginine, a fundamental amino acid, plays a pivotal role in the urea cycle, serving as a precursor for protein synthesis, polyamine production, creatine synthesis, and nitric oxide (NO) biosynthesis. Arginine deprivation has been demonstrated to have a direct pro-survival effect on myeloma cells, with potential therapeutic implications. Renal failure is a frequent clinical feature in MM patients. Creatinine, a key end product of arginine and proline metabolism, is transported to the kidneys via blood plasma, and serum creatinine is commonly used as an indicator of renal function. Previous studies have reported an obvious up-regulation of serum creatinine and arginine levels in MM patients compared with healthy controls [22, 25, 49], which is consistent with our findings. The elevated levels of creatinine in the plasma of MM patients may be attributed to impaired renal function during the progression of MM, thereby impeding the elimination of toxins [50].
Hydrovalerylcarnitine, butyrylcarnitine, L-acetylcarnitine, and L-carnitine, which play key roles in thermogenesis and fatty acid oxidation (FAO), were significantly elevated in MM patients compared with healthy controls. Acylcarnitines play a primary role in the FAO process within mitochondria, converting fatty acids into energy. When the body requires energy, fatty acids undergo β-oxidation reactions, breaking down into shorter groups and eventually transforming into acetyl-CoA to enter the tricarboxylic acid cycle, generating substantial energy for the cell. Carnitine and acetylcarnitine have been recognized as novel biomarkers for active diagnosis, relapse, and mediators of disease-associated pathologies in MM [51]. Additionally, carnitine may enhance plasma cell immunoglobulin (Ig) secretion, promoting B lymphocytes to differentiate into plasma cells and participate in antibody-mediated immune responses [52, 53]. Therefore, the increased levels of plasma carnitine and, to a greater extent, acetylcarnitine and hydrovalerylcarnitine in MM patients could entail increased lipid oxidation in highly metabolically active myeloma cells.
In conclusion, a deeper understanding of the metabolic profiles of MM could aid in identifying cases resistant to specific agents, preventing repetitive errors and cumulative toxicity, and exploring new experimental strategies for these cohorts.
Despite these insights, the study has several limitations. While we summarized differentially abundant metabolites and explored their value in MM, more functional validations in vivo and animal models are necessary. Additionally, validation and further investigation in a larger, independent cohort are warranted to better comprehend the mechanisms of MM. Further research is essential to support these results and verify the underlying biological functions of key amino acid metabolites through large-scale and mechanistic studies.
{kind=link}