Soil properties have a strong directly effect on plant functional traits of Dasiphora fruticosa on alpine region
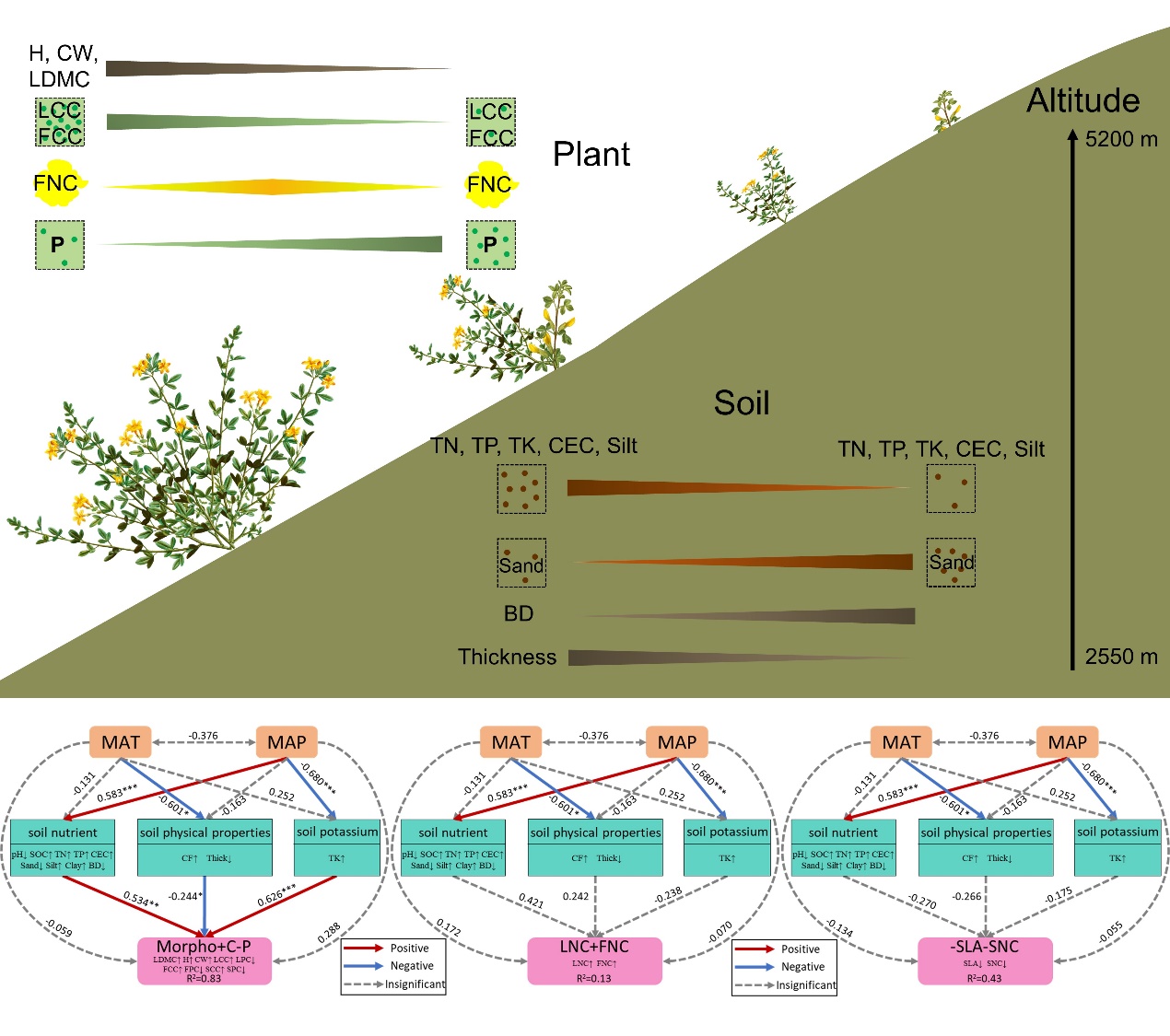
Soil properties directly and significantly affected the morphological traits, carbon and phosphorus content in organs of above-ground parts of Dasiphora fruticosa (Figs. 5 and 6a), which is consistent with other research of Dasiphora fruticosa (Qin et al. 2022). Soil properties filter out the plant functional traits of Dasiphora fruticosa directly on alpine region.
As altitude increases, TN, TP, TK, CEC, Silt and Thick of soils significantly decreased, while Sand and BD significantly increased (Table S2), and Dasiphora fruticosa tended to be shorter and smaller (Fig. 2a and b). Crown width and plant height decreased significantly along altitude and had the largest coefficient of variation (Figs. 2 and 3), indicating significant variations with environment and adaptation to changes in soil properties especially soil texture (Figs. 4, 5a and 6a). Soil texture determines soil aeration, water permeability, water retention, and fertilizer retention, all of which can impact the plant's access to water, nutrients, and oxygen (Wu and Zhao 2019). This, in turn, affects the plant's ability to develop a strong root system, ultimately influencing the overall shape and growth of the plant.
The influence of soil thickness on plant growth was mainly reflected in the supply of water and nutrients. The thicker the soil is, the larger the space is, and the plant roots could penetrate deeper into the soil to obtain more water and nutrients (Mcconnaughay and Bazzaz 1991). Research on Jatropha curcas found that when the soil thickness was larger, the individual height was taller, and the vertical space resources were relatively fully utilized (He et al. 2010b).
Leaf dry matter content was mainly influenced by soil texture (mainly sand, silt content, and soil bulk density) in this study (Fig. 4). In a study on the effect of soil texture on dry matter accumulation of Medicago sativa, it was found that the total dry matter accumulation of Medicago sativa was greater in loam clay soils than in slit loam and clay soils, suggesting that soil texture affects leaf dry matter content by affecting water and nutrient uptake by the plant (Zhao et al. 2007). LDMC is an important indicator of plant photosynthesis and biomass production (Cornelissen et al. 2003). Soil texture indirectly affects plant photosynthetic efficiency and biomass accumulation by influencing water and nutrient availability (Wu and Zhao 2019). For example, water stress reduces the rate of photosynthesis in plants and decreases the accumulation of leaf dry matter. The efficient supply of nutrients is the basis for photosynthesis and plant growth.
P content in organs of Dasiphora fruticosa was significantly affected by soil properties, especially related to soil texture (Figs. 4, 5a and 6a). In addition, P uptake by plant is indeed strongly influenced by soil texture. Soil texture determines the size, shape, and distribution of soil particles, and these factors directly affect soil porosity, infiltration rate, and water stability, which in turn affect phosphorus effectiveness and plant availability. The adsorption and desorption characteristics of phosphorus differ in soils with different textures (Chen et al. 2010; Gu et al. 2019). Under the same vegetation, loamy soils have higher C, N, and P than sandy soils (Huang et al. 2023). Soils with a high content of clay particles have a larger specific surface area and a higher cation exchange capacity, which enables them to adsorb more phosphorus. However, this adsorption may also result in the immobilization of phosphorus in the soil, with less phosphorus available to plants. In contrast, phosphorus immobilization is weaker in sandy soils, but plant uptake of phosphorus may be limited due to their lower water and fertilizer holding capacity.
As for the effect of soil nutrients, N is considered to be the main limiting factor for plant growth in many terrestrial ecosystems and one of the most important factors affecting the primary productivity (Jones et al. 2005; Luo et al. 2007; Vitousek 1982; Xing et al. 2000). The organic matter content, pH, structure and microbial activity of different soil will affect the effectiveness of nitrogen and the ability of plants to absorb nitrogen (Birch 1958; Henry and Raper 1989; Luo et al. 2007). With weak correlation to soil nutrients, nitrogen content can be stable, thereby plants exhibit stronger nitrogen retention capacity, particularly under stressful environmental conditions such as low temperature, drought and poor nutrient (He et al. 2010a; Hong et al. 2015; Zhao et al. 2017). The nutrients required for plant growth are not only dependent on the absorption of roots and leaves from soil and atmosphere, but also through the internal redistribution of nutrients in plants (Aerts 1996; Aerts and Chapin 2000; Killingbeck 1986; Xing et al. 2000). Leaf N and C contents correlated well with soil properties (Fig. 4). The poorer the soil (i.e., low in SOC, TN, TP, etc.), the lower the C and N contents in leaves and reproductive organs (Fig. 4). Overall the C and N contents in organs of Dasiphora fruticosa were less variable and maintained relatively stable contents (Figs. 2 and 3). Whereas, the relationship between P content in organs of Dasiphora fruticosa and some soil nutrients (such as SOC, TN, and TK) was not significant (Fig. 4). Qin et al. (2022) found that for leaf stoichiometric traits of Dasiphora fruticosa, P content in leaves had no significant relationship with altitude and soil properties, including SOC, TP and pH. However, leaf P content of 753 terrestrial plant species in China increased significantly with increasing soil P content (Han et al. 2005). In alpine, the stress of hydrothermal factors is greater than that of nutrients (Körner 2003; Körner et al. 1989), so that the P content in plant organs is more correlated with the soil physical properties and less correlated with the chemical properties. In addition, we only had total amounts of N, P, and K in soil, and did not measure available N, P, and K. Whether plant functional traits have a more direct response to available N, P, and K needs further study.
The change of morphological traits of Dasiphora fruticosa with environmental factors is its overall adaptation to climate, soil, altitude and other natural conditions (He 2017). It is not only affected by soil properties, but also may be an adaptive response to other environmental factors such as air temperature, precipitation, high ultraviolet radiation, low air pressure, and strong winds (Chen et al. 2015; Gao and Liu 2018; Gong et al. 2019; Körner 2003; Pan et al. 2009). The influence of mean annual temperature on the functional traits of Dasiphora fruticosa was not significant, while the mean annual precipitation had a significant effect, primarily through its indirect impact on soil chemical properties, affecting the functional traits of Dasiphora fruticosa (Figs. 5 and 6). It verified the importance of water availability in regulating the variation of plant functional traits of Dasiphora fruticosa in Qinghai-Tibet Plateau, as precipitation regulates the mobilization of soil nutrients (Müller et al. 2017; Zhao et al. 2017). Some traits that are more sensitive to temperature, such as photosynthetic resource utilization efficiency and root nutrient content (Wei et al. 2023), were not measured in this study and need further research in the future.
Dasiphora fruticosa adapts to diverse habitats by adjusting its morphological traits and phosphorus content in organs
In alpine regions, plants will undergo a series of changes to adapt to harsh environment. Among plant functional traits, morphological traits change more significantly with environment factors (Reich et al. 1999; Wright et al. 2001), which is also consistent with our findings. In this study, with the increase of altitude, plant height and crown width of Dasiphora fruticosa decreased significantly. There are similar response for other species, such as Picea abies and Larix decidua in subalpine areas (Li et al. 2003; Oleksyn et al. 1998). Mao et al. (2018) found that plant height of 4295 species of angiosperms including herbaceous plants, shrubs, and trees in Qinghai-Tibet Plateau decreased significantly along altitude. The height increment of Fagus sylvatica, Acer pseudoplatanus, Tilia platyphyllos, and Fraxinus excelsior in North-Hesse in Germany all decreased significantly along altitude (Hölscher et al. 2002). Plants grow small to adapt poor hydrothermal conditions in alpine area, which has been widely recognized in the ecological adaptation mechanism of alpine plants (Körner 2003; Körner et al. 1989; Loehle 1998).
Over a broad climatic gradient, specific leaf area and leaf dry matter content, representing leaf photosynthetic capacity, showed a trade-off relationship (Wright et al. 2004). With the increase of LDMC, leaf water content decreased, tissue density increased, so SLA decreased (Wilson et al. 1999). With the increase of altitude, temperature decreases, SLA decreases and LDMC increases (Dunbar-Co et al. 2009; Scheepens et al. 2010). This is because the colder and drier the environment, the thicker the leaves become, which increases mesophyll cell density (Zhang and Luo 2004). This allows plants to make full use of light energy, thereby to increase photosynthetic carbon fixation, and to strengthen the protective effect of leaves on high light and low temperature (Song et al. 2011). However, this study found that LDMC was significantly decreased along altitude while SLA had no significant correlation with altitude (Fig. 2), indicating that they were decoupled, which is consistent with some small-scale studies (Dolezal et al. 2019; Pan et al. 2023). Li (1996) anatomized Dasiphora fruticosa at different altitudes on the Qinghai-Tibet Plateau and found that the epidermal cells on the leaves increased along altitude, and the cells were bulliform or chimeric. The presence of bulliform cells increased the saturated fresh weight of leaves and decreased LDMC. Bulliform cells are a beneficial variant of plants in the process of natural selection, which can enhance the drought and cold resistance of plants. Additionally, SLA and LDMC were changed relatively less compared to plant height and crown width (Fig. 3), indicating the capacity of Dasiphora fruticosa to keep stable leaf photosynthesis to a wide altitude range (Odum and Barrett 2004; Tang et al. 2016; Wilson et al. 1999), which may be one of the reasons for the broad ecological amplitude of Dasiphora fruticosa.
As for stoichiometric traits, the increase of P content along altitude could be due to the fact that enzymes involved in physiological processes, such as RuBPCase (Ribulose-1,5-bisphosphate carboxylase/oxygenase) in photosynthesis, require the involvement of phosphorus, and P is a critical component of ATP (adenosine triphosphate), which is the primary energy carrier in plants (Buchanan et al. 2000; Wu et al. 2004; Ye 2010). To cope with the chill stresses in alpine, plants may need to increase their enzymatic activity to maintain their physiological functions, including photosynthesis and respiration (Liu et al. 2000; Pan et al. 2009). In addition, as altitude increases, higher leaf P content is required to maintain leaf activity and the majority of absorbed phosphorus by woody plants is used for constructing defense structures and structural materials (Onoda et al. 2004; Xie et al. 2016).
Interestingly, nitrogen content in leaves and stems of Dasiphora fruticosa was not significantly correlated with altitude (Fig. 2h and j), and coefficient of variation of nitrogen content in organs was small (Fig. 3), indicating that N content in Dasiphora fruticosa was stable. The reasons why nitrogen content in Dasiphora fruticosa remained stable are complex. Leaf N content is crucial for maintaining the stability of ecophysiological traits of plant and may involve the homeostasis mechanism in plant (Güsewell and Koerselman 2002; Sterner and Elser 2002). Organisms have the ability to maintain the relative stability of their own chemical elements in a changing environment, that is, the homeostasis mechanism (Hong et al. 2013; Kooijman 1995).
In general, changing traits were more affected by environmental factors (Li et al. 2013; Li et al. 2021; Wang and Zhou 2017). The inert response of stoichiometric traits in Dasiphora fruticosa played an important role in fully utilizing resources in extreme habitats. At different altitudes, the main regulation strategy of Dasiphora fruticosa was to adjust morphological traits and phosphorus content in organs, and maintain the stability of specific leaf area and nitrogen content to maintain the stability of photosynthesis and respiration to adapt the changing environment.
{kind=link}