This large, population-based cohort study showed a robust association between plasma GDF-15 levels and all-cause mortality. GDF-15 was also observed to be associated with coronary events, ischemic stroke and CVD mortality, but at least in part, the associations were likely to be confounded by pre-existing CV dysfunction or by the competing risk of death.
In line with previous studies [13, 17–19], the present study observed positive correlations between GDF-15 and traditional CV risk factors, including age, waist circumference, smoking, glucose, SBP, LDL and hsCRP. Since GDF-15 is an anti-inflammatory cytokine by nature [20] and all of these CV risk factors are related to systemic inflammation, the correlations could at least be partly explained by a compensatory mechanism of GDF-15 in response to inflammation. Of note, GDF-15 correlated strongly with NT-proBNP [17, 19], a commonly used marker of left ventricular dysfunction in clinical practice. Under pressure or volume overload, NT-proBNP is produced on cardiomyocytes to cope with LV hypertrophy and fibrosis due to myocardial stretch [21]. Similar to NT-proBNP, the production of GDF-15 by cardiomyocytes is stimulated in response to CV injury [3, 22, 23]. Thus, individuals with simultaneously elevated GDF-15 and NT-proBNP in this study represented a group with suboptimal CV health even in the absence of overt cardiac pathology.
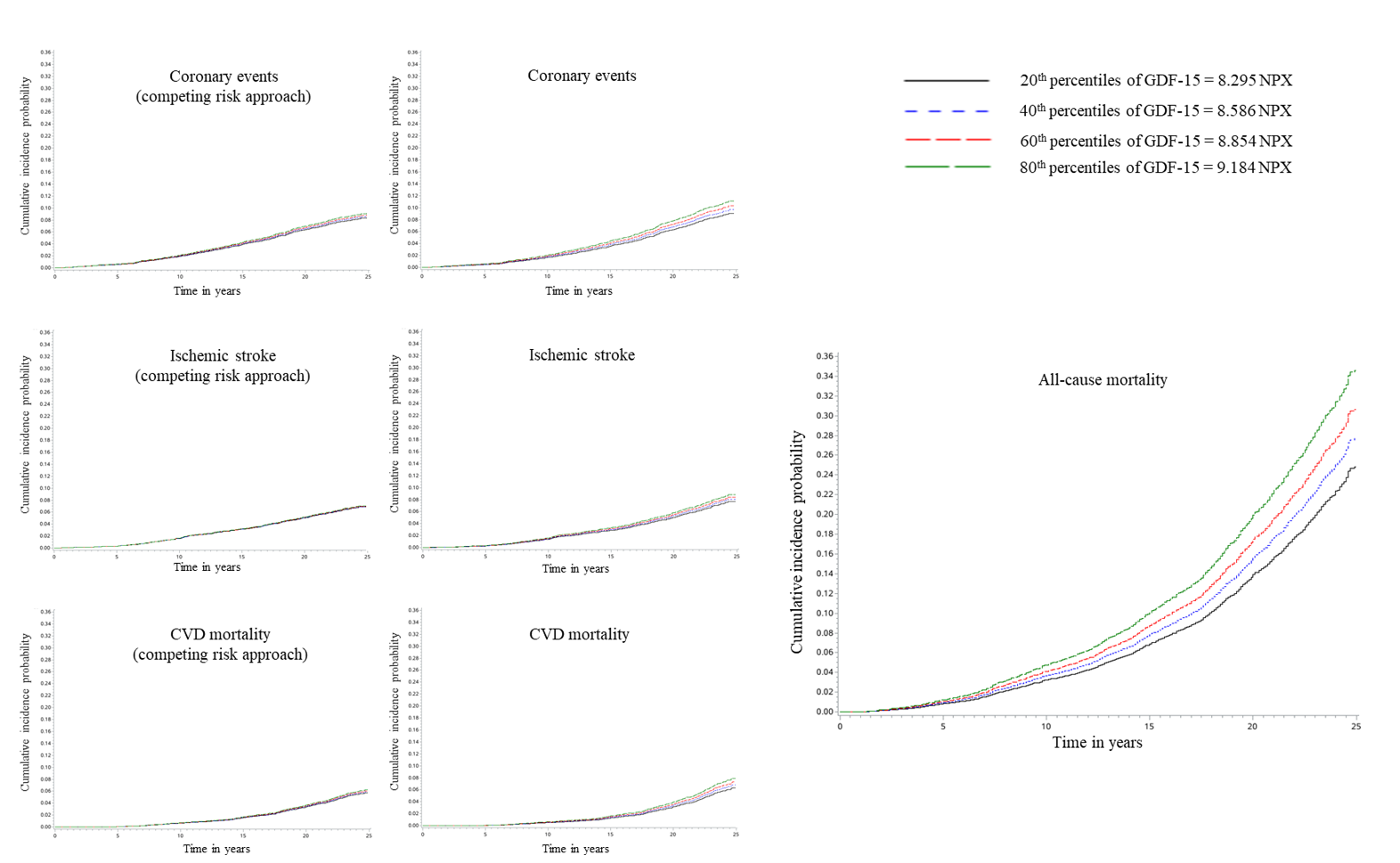
In longitudinal analyses, GDF-15 was found to be associated with incidence of coronary events, ischemic stroke, CVD mortality and all-cause mortality before correction for the confounding from NT-proBNP. This observation was in agreement with previous studies, supporting an increased risk of a wide range of CVDs in community-dwelling individuals with elevated GDF-15 levels [6, 7, 17, 19]. Our data, along with evidence from another Swedish cohort [24], suggested that the association between GDF-15 and ischemic stroke was confounded by NT-proBNP. Whereas additional adjustment for NT-proBNP greatly affected all associations with cardiovascular outcomes, its impact on the association between GDF-15 and all-cause mortality was relatively small. Previous cohort studies have consistently shown that GDF-15 and NT-proBNP are prognostic markers for adverse CV outcomes and mortality among asymptomatic subjects [6, 7, 19]. When comparisons were made between these two biomarkers, Pareek et al. [6] demonstrated that, as compared to NT-proBNP, GDF-15 showed stronger association with composite outcomes including all-cause mortality, but weaker association with incident heart failure or CVD mortality (sample size 1,324, aged 66 years, followed up for 8.6 years). Ho et al. [7] showed that GDF-15 is a better predictor of atherosclerotic CVD and all-cause mortality but a weaker predictor of CVD death than NT-proBNP (sample size 3,523, aged 62 years, followed up for 14.3 years). Similarly, Daniels et al. [19] reported that GDF-15 was a stronger predictor of all-cause mortality while NT-proBNP was a stronger predictor of CVD mortality (sample size 1,391, aged 70 years, followed up for 11 years). In addition, GDF-15 but not NT-proBNP was independently associated with non-CV death and cancer death [19]. Therefore, it seems that whereas GDF-15 might not be a better marker than NT-proBNP in a cardiovascular context, it is a powerful marker for all-cause mortality.
Noteworthy, although almost all relevant studies were conducted on an elderly population and most of them were with a long-term follow-up, hardly any study has carefully evaluated the death as a competing risk. Emphasis has been made that the competing risk of death is an important consideration in studies evaluating the long-term association among older adults, ignoring of which might lead to substantially overestimated associations [8, 9]. Kaplan-Meier estimates and traditional Cox proportional hazards regression models assume that subjects are censored from causes unrelated to the primary outcome. Therefore, those who died from other causes are still considered to be at risk of the primary outcomes at the point of censoring and are thought to have the same distribution of time-to-event as incident cases. However, such subjects cannot possibly develop the primary outcomes, thus ignoring that the competing risk of mortality could exaggerate the HRs for the primary outcomes, particularly for subjects with high mortality, e.g. older adults with long-term follow-up [8, 9]. Our study added important information on the competing risk of death and indicates that the associations of GDF-15 with coronary events and CVD mortality may be markedly overestimated if this competing risk is ignored, especially given that high GDF-15 contributed largely to mortality. As a result, only all-cause mortality remained significantly related to GDF-15 in this study. The association between GDF-15 and all-cause mortality was strengthened in subjects with higher baseline hsCRP, which may be due to an enhanced secretion and function of GDF-15 in pro-inflammatory conditions.
Of note, accumulating evidence has indicated that GDF-15 has a role beyond inflammation [1, 17]. It has multiple roles in, for example, oxidative stress, tumorigenesis, strain and ischemia, and its elevation is associated with a wide range of diseases involving different systems [1, 17]. Therefore, it is speculated that GDF-15 concentration reflects overall health status and thus correlates with all-cause mortality. Elevated GDF-15 levels mirror cellular stress and senescence, which might also explain the association between GDF-15 and life span. As a downstream target of p53, GDF15 gene is expressed via a p53-dependent pathway in cellular response to a broad range of stress [25, 26]. During this process, p53 mediate cellular senescence and apoptosis by functioning as a transcription factor [26], and thus plays a role in regulating longevity [26]. At structural levels, p53 directly influences mitochondrial respiration and impairs clearance of defective mitochondria [27, 28], leading to mitochondrial dysfunction, a well-known driver of biologic ageing [29]. Consistently, emerging studies have proposed GDF-15 as a specific biomarker of mitochondrial disorder [30, 31]. In further support of GDF-15 as a marker of ageing, GDF-15 has recently been identified as a soluble senescence-associated secretory protein and a key of cellular senescence [32]. Nevertheless, it remains largely obscure whether and exactly how GDF-15 itself promotes ageing.
The study is based on a large number of subjects from the general population, with a long-term follow-up and sufficient number of cases. Less than 1% was loss to follow-up [10]. CV endpoints were retrieved from validated national and local registers [14, 15]. However, there are some limitations that might have affected our study. High-risk individuals are less likely to participate in population-based studies. Thus, the proportion of individuals with high risk could be underestimated. Information on biomarkers was available for a sub-cohort, MDC-CV, of the whole MDC cohort. Nevertheless, the sub-cohort was randomly selected and should be regarded as representative of the whole MDC cohort in terms of age, sex, biological, lifestyle and socio-economic characteristics [33]. Furthermore, the risk factors used for adjustments in the study are well-known CV risk factors but still we cannot exclude the possibility of residual confounding. Information about incident coronary events and ischemic stroke was retrieved from local and national hospital registers. The validity of this information has been confirmed in many previous studies [14, 15].
{kind=link}