Physiological responses to salt stress
We examined leaf relative water content (RWC), superoxide dismutase (SOD) and catalase (CAT) activities, proline (PRO) and soluble sugar (SS) contents in alfalfa plants with active nodules (NA), inactive nodules (NI) and without nodules (NN) under different extents of salt stress (Fig. 1). RWC of alfalfa leaves decreased with the time of salt stress and was higher in inoculated plants (NA and NI) compared to NN 11 d after salt stress treatment (Fig. 1A). The activities of SOD and CAT were found to be the highest in NA shoots under salt stress, and the difference was more noticeable on day 10 among NA, NI, and NN plants (Fig. 1B and 1C). PRO and SS contents in NA and NI shoots were higher than those in NN after salt stress (Fig. 1D and 1E). A greater PRO content in NA shoots was observed on day 10 after stress compared to NI shoots. NA and NI plants also displayed higher SS contents on day 10 after salt stress compared to NN plants. Overall, NA plants show different physiological responses to salt stress treatment on day 10 compared to NI and NN plants, and the samples harvested on day 10 after salt stress were studied for difference in protein profiles among NA, NI and NN plants.
Protein identification
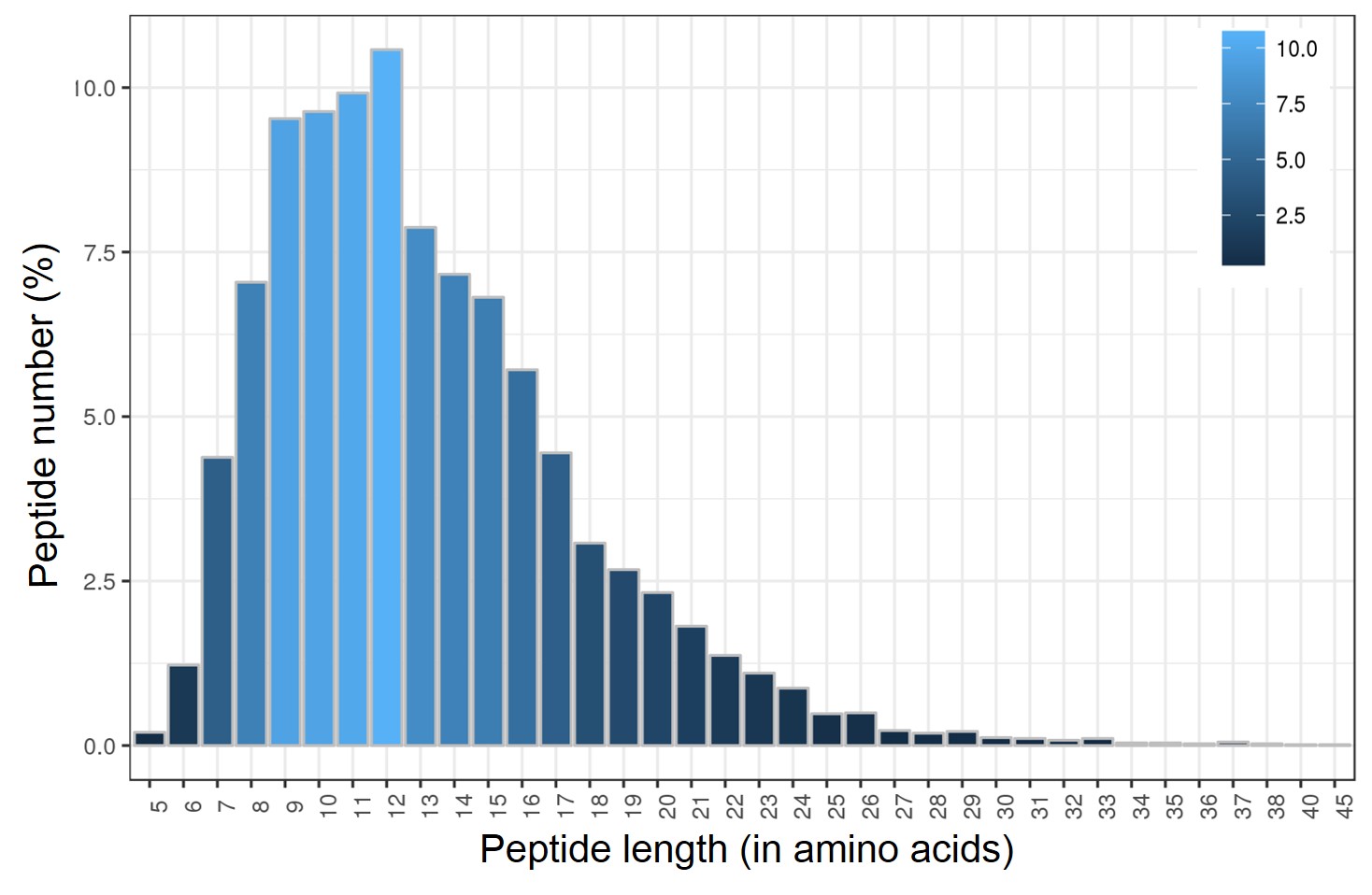
An iTRAQ-based quantitative proteomic approach was used in this study to investigate the molecular links between root nodules and alfalfa salt tolerance. With two biological replications, a total of 396,885 spectra were obtained, among which 33,896 spectra were matched to the reference genomes M. truncatula protein database (ftp://ftp.jcvi.org/pub/data/m_truncatula/Mt4.0/Annotation/ Mt4.0v1/), representing 29,981 unique spectra. A total of 9,240 peptides were identified, representing 8,717 unique peptides which can be assembled into 3,274 proteins (Fig. 2A). Majority of the peptides (85%) are 7– 17 amino acid residues long (Fig. S1). Over 50% of the 3,274 proteins identified contain at least two peptides (Fig. 2B).
The protein mass distribution was presented in Figure 2C. Proteins with 20–30 kDa were the most abundant, followed by proteins with 30-40 and kDa 10–20 kDa. Our results showed that iTRAQ was able to identify proteins with a wide range of molecular mass. Forty one percent of the identified proteins had more than 10% of the peptide coverage, and 19% have more than 20% of the peptide coverage (Fig. 2D).
Repeatability analysis
In this iTRAQ-based proteomics, the distribution of the coefficient of variation (CV) showed that our data had a relatively high repeatability with two biological replicates. The mean CV value is 0.17 (S_NA:CK_NA), 0.17 (S_NI:CK_NI), and 0.19 (S_NN:CK_NN) for three comparative groups. When the level of variation was < 20%, the proportion of the number of proteins was 0.70, 0.69, and 0.68 for S_NA:CK_NA, S_NI:CK_NI, and S_NN:CK_NN, respectively (Fig. 3).
Overall differentially expressed proteins in alfalfa tissues
A total of 3,274 non-redundant proteins in alfalfa shoots were identified (Supplementary Table S2). These proteins were used to determine the significant changes in abundance during salinity treatments. With a 1.2-fold change cutoff and p-value less than 0.05, 607 proteins are differentially expressed in the shoots when comparing the salt stress condition with normal growth conditions in each treatment (Supplementary Table S3). Among the 607 proteins, 408 were up-regulated and 322 were down-regulated (Fig. 4). NN, NI, and NA shared 109 up-regulated and 61 down-regulated proteins. The numbers of proteins that are up-regulated specifically in NN, NI, and NA are 54, 72 and 53, respectively; and the numbers of proteins that are down-regulated specifically in NN, NI, and NA are 47, 52, and 55. Pairwise comparison showed that NI and NA shared similar changes when each of NI or NA was compared to NN. NI and NA shared 79 up-regulated and 71 down-regulated proteins, while NN only shared 23 and 18 up-regulated proteins with NI and NA and 25 and 11 down-regulated proteins with NI and NA, respectively.
Functional classification of differentially expressed proteins
Clusters of orthologous groups of proteins (COG) analysis was performed to predict the functions and phylogenetic annotations of differentially expressed proteins in NA, NI and NN tissues under salt stress (Fig. 5). NN, NI and NA all showed a higher up-regulation of proteins involved in “energy production and conversion (C)”; “amino acid transport and metabolism (E)”, “carbohydrate transport and metabolism (G)”, “translation, ribosomal structures and biogenesis (J)”; “posttranslational modification, protein turnover, chaperones (O)” and “general function production only (R)”. NA and NI generally showed similar but higher levels of changes when compared to NN in C and E functional categories. The most highly down-regulated functional categories are the same for NN, NI and NA, i.e. “translation, ribosomal structures and biogenesis (J)”; “posttranslational modification, protein turnover, chaperones (O)” and “general function production only (R)”. Salt stress appeared to have little effect on “cell motility (N)”. COG analysis for NA, NI and NN plants showed that the three most down-regulated functional categories are among the six most up-regulation functional categories, and two of them, “translation, ribosomal structure, and biogenesis” and “posttranslational modification, protein turnover, chaperons”, are involved in protein synthesis, modification or degradation, suggesting plants actively regulated protein quantity and activity during salt stress response, resulting in a large increase of proteins in “energy production and conversion”, “amino acid transport and metabolism”, and “carbohydrate transport and metabolism”.
Pathway analysis for differentially expressed proteins with MapMan
To better understand how NA, NI and NN plants respond to salt stress differently, MapMan was used to compare the proteins involved in specific processes. As indicated in Figure 6, seven processes were upregulated in NA, NI and NN plants, and among them four processes are highly over-represented, and they are all related to photosynthesis. Three upregulated processes common to NA, NI and NN plants are under-represented and belong to “not assigned” categories (Fig. 6). Interestingly, the only down-regulated process common to NA, NI, and NN plant also belongs to “not assigned unknown” category.
MapMan analysis revealed 5 processes were upregulated and 12 were downregulated only in NA under salinity based on the over-represented proteins (Fig. 6). Four of the up-regulated processes are related to cell wall modification. They include “cell wall degradation”, “miscellaneous”, “miscellaneous gluco-, galaco- and mannosidases” and “miscellaneous peroxidase”. Among the 12 downregulated processes, 8 of them are related to protein metabolism, including two for protein degradation, four for protein synthesis and two for protein targeting/secreting. NA plants also showed downregulation of ethylene responsive proteins, receptor kinases and calcium signaling proteins.
Both NA and NI showed significant up-regulation of proteins involved in photosynthesis and respiration, including proteins in light reaction of PSI, in the Calvin cycle, such as fructose-bisphosphate adolase, in photorespiration, and in TCA cycle. NA and NI plants also showed a stronger up-regulation of proteins involved in biodegradation of xenobiotics and porin transport.
{kind=link}