3.1. Hunger Induction Affects Exploratory Behavior in Zebrafish
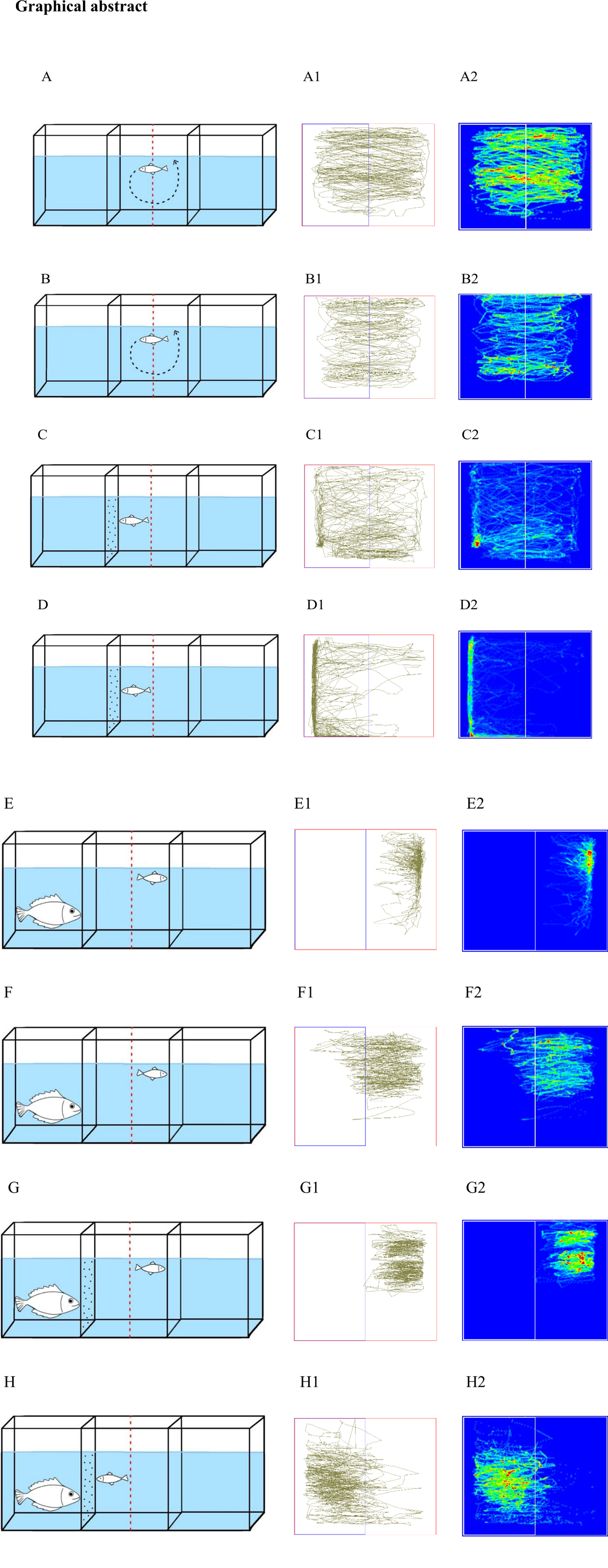
To investigate the effects of hunger on zebrafish exploratory behavior, we used wild-type zebrafish deprived of food for 6 days. We adapted a protocol used to induce starvation from (Nakajo et al., 2020). Figure. 1 represents the effect of food deprivation on the exploratory behavior of control (CNTRL) and hungry (HGRY) zebrafish. The result revealed that the motor activity parameters like, total distance travelled (Fig. 1A, cohen’s d = -1.84 [95.0%CI -2.56, -1.01], two-sided permutation t-test p = 0.0001) and total speed (Fig. 1C; cohen’s d = -1.84 [95.0%CI -2.55, -1.13], two-sided permutation t-test p = 0.0001) were decreased in the starved fish when compared to control, potentially due to the depletion of energy reserves, and reduced motivation to explore or engage in movement. Which limit the fish capacity to sustain normal locomotor activity, reflecting an adaptive strategy to conserve energy during periods of food scarcity (Wang et al., 2006).
While total resting time (Fig. 1B, cohen’s d = 1.29 [95.0%CI 0.897, 1.7], two-sided permutation t-test p = 0.0001) was higher in the HGRY group, the number of zone transitions (Fig. 1D, cohen’s d = -0.457 [95.0%CI -1.24, 0.22], two-sided permutation t-test p = 0.1990) was non-significant, though the test group made fewer transitions compared to the control group. This suggests that food deprivation may significantly increase the amount of resting time, it does not significantly impact the frequency of transitions between zones. This could indicate that although the HGRY group is more sedentary overall, their exploratory behavior remains relatively stable, possibly due to the prioritization of energy conservation over increased activity. Together, these energy state-dependent behavioral adaptations enhance the probability of finding food while reducing the energy expenditure (Smith & Grueter, 2022).
3.2. Visual Stimuli Trigger Prey Capture-related Responses in Hungry fish.
Hungry zebrafish exhibit a pronounced increase in object orientation and prey capture-related behavior, specifically dot biting when exposed to artificial visual stimuli (as black dots) mimicking prey, reflecting their heightened predatory drive. Hunger, as shown by earlier studies (Semmelhack et al., 2014; Bianco & Engert, 2015), alters brain circuits linked to prey detection, increasing an organism's sensitivity to visual cues that resemble prey.
The behavior of the CNTRL+D group exhibited a significant increase in the time spent within the stimulus zone (Fig. 2A&B p = 0.0181) and a markedly higher frequency of bites on black dots compared to controls (Fig. 4B, p = 0.0001). Figure 4.A displays the variability in pairwise relationships of dot bite frequencies highlights the differential impact of the experimental conditions on prey capture behavior across different groups. This suggests that the normal-fed zebrafish also perceive small black dots as prey and show hunting behavior in the form of bites. However, no significant difference was observed in latency to enter the stimulus zone (Fig. 2C&D, p = 0.08175), the number of zone transitions (Fig. 3B, p = 0.4407), or entries into the stimulus zone (Fig. 2E&F, p = 0.4250).
In the comparison between HGRY and HGRY+D groups, the HGRY+D cohort demonstrated significant increase in time spent in the stimulus zone (Fig. 2B, p = 0.0001) and a higher number of dot bites (Fig. 4B, p = 0.0001). This indicates that hungry zebrafish not only recognized the black dots as prey but also spent more time in the stimulus area and displayed heightened biting behavior. Despite these observations, the HGRY+D group showed reduced zone transitions (Fig. 3B) and fewer entries into the stimulus zone (Fig. 2E&F, p = 0.0001), suggesting that while the fish focused their interaction more with the stimulus (prey), their overall exploratory behavior was less dynamic. Figure 2D represents individual density plots that visualize the distribution of latency for zebrafish to enter the stimulus zone, showing the variation in response across different experimental conditions. Additionally, the latency to enter the stimulus zone was not significantly different (Fig. 2D, p = 0.0585), indicating that while the HGRY+D fish showed a stronger predatory response, their initial reaction time to the stimulus was comparable to that of the HGRY group. These results highlight, the addition of prey (black dots) enhances prey-directed behavior in food deprived zebrafish and alters their exploratory behavior pattern.
CNTRL+D and HGRY+D groups revealed that hungry zebrafish (HGRY+D) spent significantly more time in the stimulus zone (Fig. 2B, p = 0.0001) and exhibited a higher number of bites on the black dots (Fig. 4B, p = 0.0001) compared to their well-fed counterparts. This suggests a notable shift in the activity pattern by nutritional status, with hungry fish demonstrating a stronger predatory response to the black dots, interpreting them as prey and increased their engagement. In contrast, HGRY+D fish showed significantly fewer zone transitions (Fig. 3B, cohen’s d = -2.62 [95.0%CI -3.44, -1.61]) and reduced entries to the stimulus zone (Fig. 4E&F, p = 0.0001) relative to CNTRL+D fish. This decrease in exploratory behavior may indicate that, the hungry fish has increased time spent in the stimulus zone, which is associated with a more focused, albeit less exploratory, intense predatory strategy. Our results reveal that the orientation towards these artificial stimuli is closely linked to the motivational state of hunger, which modulates the zebrafish to exhibit predator-like behavior, including increased attention and engagement with prey-like stimuli. This finding also indicates that hunger not only enhances the fish focus on potential food sources but also influences their behavioral patterns to mimic those observed in natural predatory state. This pattern highlights a distinct shift in behavior, transform their normal behavior to a goal-oriented approach, typical of predator-prey interactions.
3.3. Intrinsic Metabolic Demand Prime the Individuals in a Challenging Environment for Risk Taking Behavior
The physiological state of food deprivation has been observed to prime zebrafish for risk-taking behavior in challenging environment, as evidenced by their altered responses to predator cues. Food-deprived zebrafish exhibit increased boldness, spending more time in proximity to predator zone, reflecting a heightened willingness to take risks compared to their control fishes. Animals receiving limited dietary treatments are more likely to exhibit high-risk behavior in a variety of situations involving predation (Moran et al., 2021).
The comparison between CNTRL and CNTRL+PDTR groups, the CNTRL+PDTR fishes exhibited a significantly longer latency to enter the stimulus zone (Fig. 2C&D, p = 0.0004). Additionally, the time spent in the stimulus zone (Fig. 2B, p = 0.0001), the number of zone transitions (Fig. 3B, p = 0.0001), and the resting time within the stimulus zone (Fig. 3A, p = 0.0151) were all significantly reduced compared to the CNTRL group. These findings suggest that the presence of a predator stimulus in the CNTRL+PDTR group induced an aversive response, leading to decreased exploratory behavior and reduced engagement with the stimulus zone. This indicates that the predator stimulus effectively altered the zebrafish behavior, making them more cautious or fear in the presence of a potential threat.
Similar to the above findings, zebrafish in the HGRY+PDTR group displayed a significantly longer latency to enter the stimulus zone (Fig. 2D, p = 0.0221). Also, the zebrafish spent less time in the stimulus zone (Fig. 2B, p = 0.0001), exhibited fewer zone transitions (Fig. 3B, p = 0.0069), and had reduced resting time within the stimulus zone (Fig. 3A, p = 0.0240) compared to the HGRY group. The observed changes reflect an avoidance behavior consistent with heightened threat perception in the presence of a predator.
In the case of CNTRL+PDTR and HGRY+PDTR groups, the HGRY+PDTR demonstrated significantly higher time spent in the stimulus zone (Fig. 2B, p = 0.0001), more zone transitions (Fig. 3B, cohen’s d = 1.27 [95.0%CI 0.405, 2.04]), and increased resting time in the stimulus zone (Fig. 3A, cohen’s d = 0.861 [95.0%CI 0.51, 1.26]). These findings suggest that hunger may drive a more aggressive or bold behavior, potentially mitigating the intensity of fear responses triggered by the predator stimulus. Despite both the CNTRL+PDTR and HGRY+PDTR groups shows aversive behavior to their sympatric predator (anabas), the HGRY+PDTR condition exhibited a significantly heightened approach behavior compared to CNTRL+PDTR, as evidenced by increased time spent in the stimulus zone, more zone transitions, and longer resting time in the stimulus zone. This behavior reflects an adaptive response where the fish weighs the immediate need for food against the potential cost of predation, demonstrating the complex interplay between internal state (like hunger) and external environmental factors (like predatory threats) in decision-making processes. The nutritional state plays a crucial role in contexts involving both direct and indirect predation risk. Similarly, in risk-taking scenarios where physical condition offers a clearer benefit, such as intraspecific contest, the nutritional state becomes a significant factor (Moran et al., 2021). This hunger-driven behavioral shift highlights a complex interaction between motivational states and risk assessment, where the need for food may override the cautionary behavior, typically seen in the presence of a predator.
3.4. Food Deprivation Reverse Aversive Behavior to an Approach Strategy
This study confirms, food deprivation can reverse typical predator avoidance to a more exploratory and approach-oriented strategy. In a predatory environment, food-deprived zebrafish were observed to approach the black dots (prey) rather than the expected fear-driven aversive response seen in well-fed individuals. Similar findings have been reported in earlier studies, where a risk-taking behavior was displayed by fishes following food deprivation (Gotceitas & Godin 1991; Pettersson & Brönmark 1993; Damsgird & Dill 1998; Krause et al., 1998).
The CNTRL+D+PDTR group exhibited a significantly longer latency to enter the stimulus zone (Fig. 2D, p = 0.0001). In addition, there was a notable decrease in the time spent in the stimulus zone (Fig. 2B, p = 0.0001), the number of entries into the stimulus zone (Fig. 2F, p = 0.0001), and the number of bites on the dots (Fig. 4B, p = 0.0001) when compared to CNTRL+D group. These results indicate that the presence of the predator stimulus induces a fear response even in satiated zebrafish, leading to reduced exploration and engagement with the stimulus. In the HGRY+D+PDTR group, both the time spent in the stimulus zone (Fig. 2B, p = 0.0001) and the number of bites on the dots (Fig. 4B, p = 0.0001) were significantly reduced when compared to the HGRY+D group. This reduction reflects an aversive response to the predator stimulus in hungry fish. Despite this, HGRY+D+PDTR zebrafish exhibited a higher number of entries to the stimulus zone (Fig. 2F, p = 0.0002), indicating a risk-taking behavior.
Comparing the CNTRL+PDTR and CNTRL+D+PDTR condition, the CNTRL+D+PDTR group showed a significant reduction in time spent in the stimulus zone (Fig. 2B, p = 0.0483), indicating an increased fear response induced by the predator stimulus. However, other parameters including latency to enter the stimulus zone (Fig. 2D, p = 0.4987), entries to the stimulus zone (Fig. 2F, p = 0.1093), resting time (Fig. 3A), and mean speed (Fig. 3C, p = 0.5968) were not statistically significant. While the HGRY+PDTR and HGRY+D+PDTR, the HGRY+D+PDTR group exhibited a significantly reduced time spent in the stimulus zone (Fig. 2B, p = 0.0001) and resting time in the stimulus zone (Fig. 3A, p = 0.0027). Conversely, the latency to enter the stimulus zone was significantly decreased in the HGRY+D+PDTR group (Fig. 2D, p = 0.0002). This indicates a perceptual shift in hungry fish exposed to both predator and prey stimuli, where prey perception and approach behavior appeared to dominate over predator-induced avoidance behavior, reflecting an increased risk-taking tendency in the hunger state.
In CNTRL+D+PDTR and HGRY+D+PDTR groups, zebrafish in the HGRY+D+PDTR group exhibited a significant increase in time spent in the stimulus zone (Fig. 2B, p = 0.0001), entries to the stimulus zone (Fig. 2F, p = 0.0001), resting time in the stimulus zone (Fig. 3A, cohen’s d = 1.08 [95.0%CI 0.821, 1.49]), and number of dot bites (Fig. 4B, p = 0.0001). Additionally, the latency to enter the stimulus zone was significantly reduced in the HGRY+D+PDTR group (Fig. 2D, p = 0.0001). These findings suggest that hunger drives zebrafish to focus more actively to the stimulus (prey), indicating a shift in the risk-taking behavior even in the presence of predator cues. To determine the frequency of prey capture behavior among the various conditions (CNTRL, HGRY, CNTRL+D, HGRY+D, HGRY+D+PDTR, and CNTRL+D+PDTR), we calculated the average pairwise correlation of the number of dot bites. The analysis revealed that the number of dot bites was negatively correlated between the HGRY+D and CNTRL+D conditions, as well as between the HGRY+D+PDTR and CNTRL+D+PDTR conditions (Fig. 4C). This indicates that as the frequency of prey capture behavior increased in one group, it decreased in the other. With the exception of CNTRL and HGRY, where the approach behavior showed a strong correlation, most paired groups exhibited relatively weak correlations. The prey capture behavior among different conditions are not uniformly consistent, reflecting variability in the extent to which hunger and predator presence influence prey capture.
The increased number of dot bites, higher entries into the stimulus zone, reduced latency, and extended resting time within the stimulus zone illustrate that food-deprived zebrafish are willing to take the risk of encountering a predator by abandoning defensive behavior in favor of securing food. The tendency of risk taking decision during foraging seems to be regulated by the feeding state (Filosa et al., 2016). This shift in behavior, characterized by an increased approach towards prey despite the potential threat, underscores how hunger can alter perception and influence decision-making process in zebrafish. Previous research by Killen et al. (2011) also revealed how intrinsic metabolic demand can affect risk-taking behavior. The observed reversal from aversion to approach in food-deprived zebrafish highlights the profound impact of physiological state on decision-making process, particularly in balancing the trade-off between avoidance and approach.
{kind=link}