All animals are associated with and are colonized by communities of microorganisms, known collectively as the microbiome. Animals obtain microbes from the environment and their behavior and life stage also have a large impact on their microbiome. Throughout their lives, animals are colonized by microbes from their surroundings [1–3]. Because microbes are acquired in various ways, composition of microbial communities depends on how they are obtained or transmitted. Studies in humans and non-human primates suggest that the surrounding environment including habitat, diet, or social group can significantly influence microbiome composition [4–6]. The composition of the gut microbiome varies across captive, urban, and rural environments in many organisms such as Ring-tailed lemurs [7], Tasmanian devils [8], deer mice [9], water dragons [10], and coyotes [11]. Host behaviors significantly impact host-microbiome dynamics. For example, communal nesting in four-toed salamanders (Hemidactylium scutatum) increases the transmission of beneficial, antifungal bacteria, enhancing hatchling survival compared to solitary nests [12, 13]. The diversity of host behaviors is further highlighted by the fact that both vertebrates and invertebrates, such as the Green iguana (Iguana iguana) and bumblebees (Bombus), engage in coprophagy, a behavior involving the consumption of feces to establish and regulate their gut microbiota [14, 15].
Most microbiome studies consider microbial communities inside or on the surface of the organism. However, many animals occupy stable burrows or construct nests [16], which may have a large impact on the microbial environment of the animals. In some cases, like social insects, nests can be considered to be part of the organism, as an extended phenotype [17, 18]. Thus, the microorganisms inside a nest can be considered as part of the microbiome of a colony of social insects. Little is known about the feedback between the microbiome of a nest and how it relates to the microbiome of its inhabitants. Furthermore, social insects are known for their intricate social organization, with a morphological division of reproductive labor, as well differentiation in the tasks that workers perform. Furthermore, ants use their nest to raise brood and store food leading to highly diverse contents within an ant colony’s nest. Therefore, social insects afford a unique opportunity to determine the role of the environment and social organization on microbiome diversity and composition.
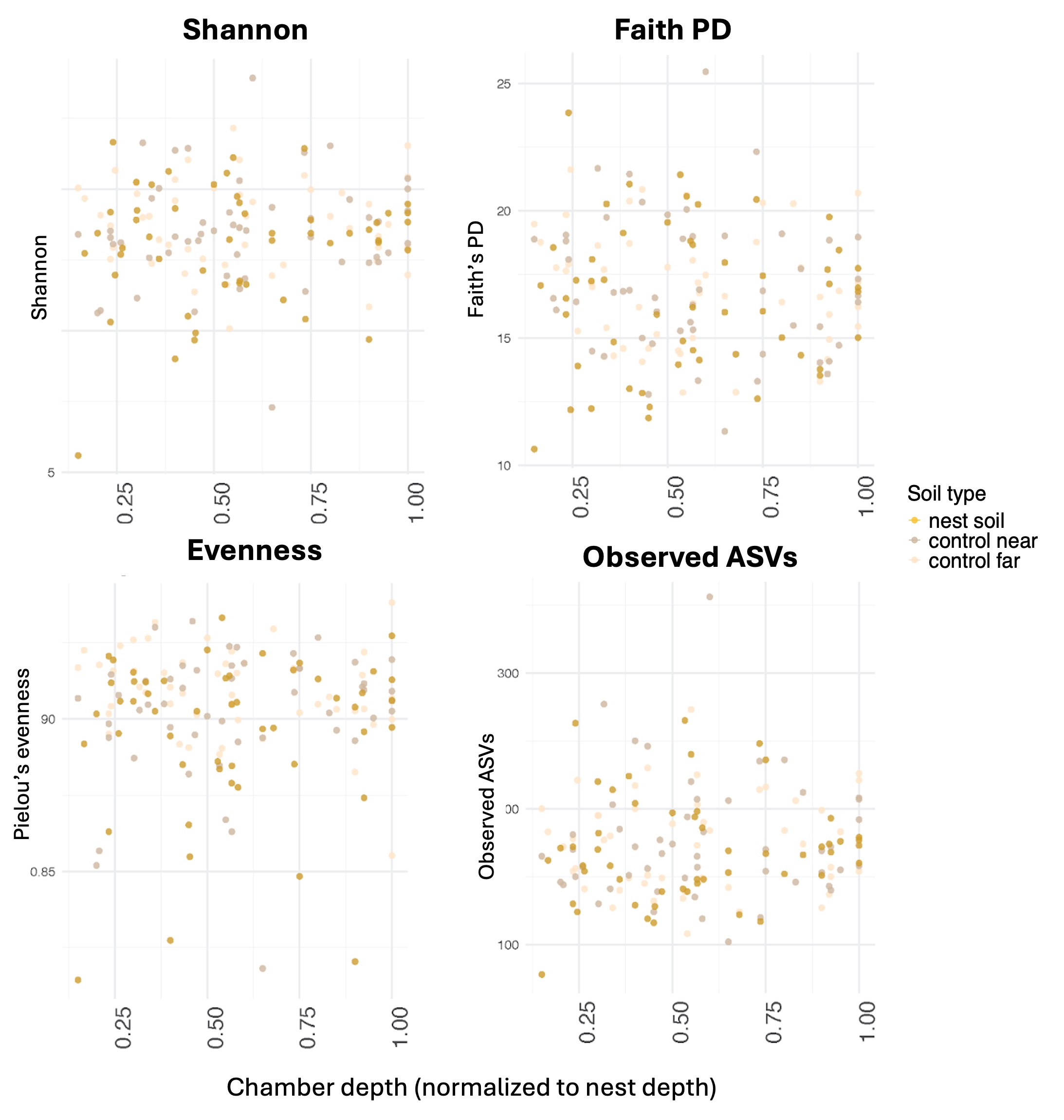
Environmentally acquired microbes tend to be ephemeral and not host-specific due to the functional redundancy of bacterial species and the changing environmental conditions that both hosts and their microorganisms are exposed to (temperature, humidity, nutrients) [19]. Indeed, the microbiome of animals is often determined by the environment in which they live. For example, when the cuticular microbiomes of two arboreal ant species were compared, the physical location of their nest was a better predictor of their microbiome composition than the species of ant [20]. Similarly, when comparing the gut microbiome of deer mice (Peromyscus manuculatus) in captive and urban populations, individuals from each environment had distinct microbial compositions. Therefore, the relative impact of the environment on the microbiome of an animal is important to consider, especially for animals whose environment is an integral component of their phenotype, such as soil-nesting ants. Microbial diversity in soils is linked to soil pH, soil organic carbon, and oxygen [21, 22]. Furthermore, microbial biomass and diversity tend to decrease with soil depth [23]. Therefore, we expect that if the microbiome of animals that live in soil is impacted by the environment, such subterranean animals will have microbiomes that mirror the soil’s, including decreased microbial diversity with depth.
The behavior of an animal and the social organization of a society can impact the microbiome of an animal and the microbiome can provide information specific to the host. For example, in spotted hyaenas, the microbiome varies with sex and age-class and is specific among individuals [24]. The social organization of social insect colonies results in individuals performing different tasks and this division of labor can influence and structure the microbiome composition of individuals within colonies. For example, honeybee workers that perform different behavioral tasks, such as foraging, or nursing, show differences in gut bacterial community composition [25]. Additionally, there are differences in the gut microbiome composition of reproductives and non-reproductive workers in termites [26, 27], honey bees [28, 29], and ants [30, 31]. When comparing the gut microbiome of chimpanzees, individuals from the same communities had a similar microbiome composition, as expected, due to their shared diets and interactions. However, individuals who were considered long-term immigrants in that community showed more distinct gut microbiome composition, suggesting that the immigrant individuals retained characteristics of their gut microbial communities for extended periods regardless of the environment [32]. Thus, in some cases the behavioral role of individuals might have a stronger impact on their microbiome than the environment in which they live.
Ants are highly social animals that shape the environment in which they live - their nest. The microbial communities of the nest are shared with the microbiome of the ant colony [20]. However, most studies of ant microbiomes have focused on the gut microbiome, showing that ant species differ in the densities of bacterial communities in the gut according to diet type [33] and that ants can benefit from their microbiome via nutrient acquisition and defense against pathogens [34, 33]. However, to our knowledge, the role of the nest in shaping the microbiome of ant colonies has only been explored in arboreal ants that occupy exiting tree cavities [35] and not in subterranean ants that construct and shape their own nest. Each nest region has a different chemical signature that reflects the individuals that occupy that area [36]. Thus, it is likely that a similar relationship between the structure of the nest and the materials inside each chamber shape the microbiome of the colony and its nest, as seen in arboreal ants [35]. Behavioral tasks occur at specific locations within a nest, such that when not foraging, foragers are found near the entrance of the nest and brood nurses are found in the center, where the brood is located [37]. This spatial division of labor can influence and structure the microbiome composition of individuals within colonies of ants. The relationship between social organization and spatial position, as well as the nest being an extended phenotype of a colony suggests that the physical environment and social organization combine to influence the microbiome of ant colonies. For example, nest chambers might differ in their microbiome composition based on the behavioral tasks performed in them. Such potential differences in chamber microbiome can be driven by the chambers’ content (e.g., the seeds or the brood) or by the ants that tend to the chamber material (Fig. 1). Furthermore, the microbiome found inside ant nests might differ from the surrounding soil. Indeed, the mounds of harvester ant colonies can have a different nutrient composition compared to the surrounding soil [38], however, it is not known if such differences permeate the inside of the nest.
Colonies of the harvester ants, Veromessor andrei, live in grasslands, where they turn and aerate the soil, redistributing nutrients and potentially creating favorable conditions for microorganisms within and around their subterranean nests [38–41]. Veromessor andrei nests provide an opportunity to examine the effects of the physical environment and social organization on the microbiome of the colony. The structure of V. andrei nests affect their collective foraging [42] most likely through the impact of the nest structure on interactions among ants [43] which are known to regulate foraging activity in other harvester ants within the colony [44–46]. Thus nest structure impacts V. andrei behavior, and can potentially further segregate behavioral tasks such as brood care and food storage. However, it is not known how nest structure and social organization combine to impact the microbiome of harvester ant colonies.
To determine the roles of the physical environment and social organization in structuring an organism's microbiome we examined the microbial communities within and around nests of V. andrei. If the physical environment influences microbial communities we predict that: 1) the microbiome of the content of the nest (like ants, seeds, brood, etc) will be similar to the surrounding soil; 2) the microbiome of the soil inside nest chambers will not differ from the microbiome in the surrounding soil; 3) microbiome diversity in nest chambers will decrease with nest depth, similarly to the relationship between depth and microbiome that can be found in soils [23] and 4) nests in different locations will have different microbial diversity because soils change their microbial composition and diversity spatially [21, 22] (Fig. 1A). If social organization influences microbial communities of ant nests we predict that: 1) nest content (like ants, seeds, brood etc) will differ in their microbiome composition according to their biological classification and will be different from the surrounding soil; 2) microbiome diversity of chamber soil will differ across chambers according to the content found in them, regardless of the surrounding soil; 3) microbiome diversity of chamber soil will differ from the microbiome of the surrounding soil and 4) microbiome composition of soil inside nest chambers will be conserved by the content of the chamber in a way that is consistent across different nests (Fig. 1B).
{kind=link}