3xTg mice displayed decision-making deficits in the 4-Choice Gambling Task.
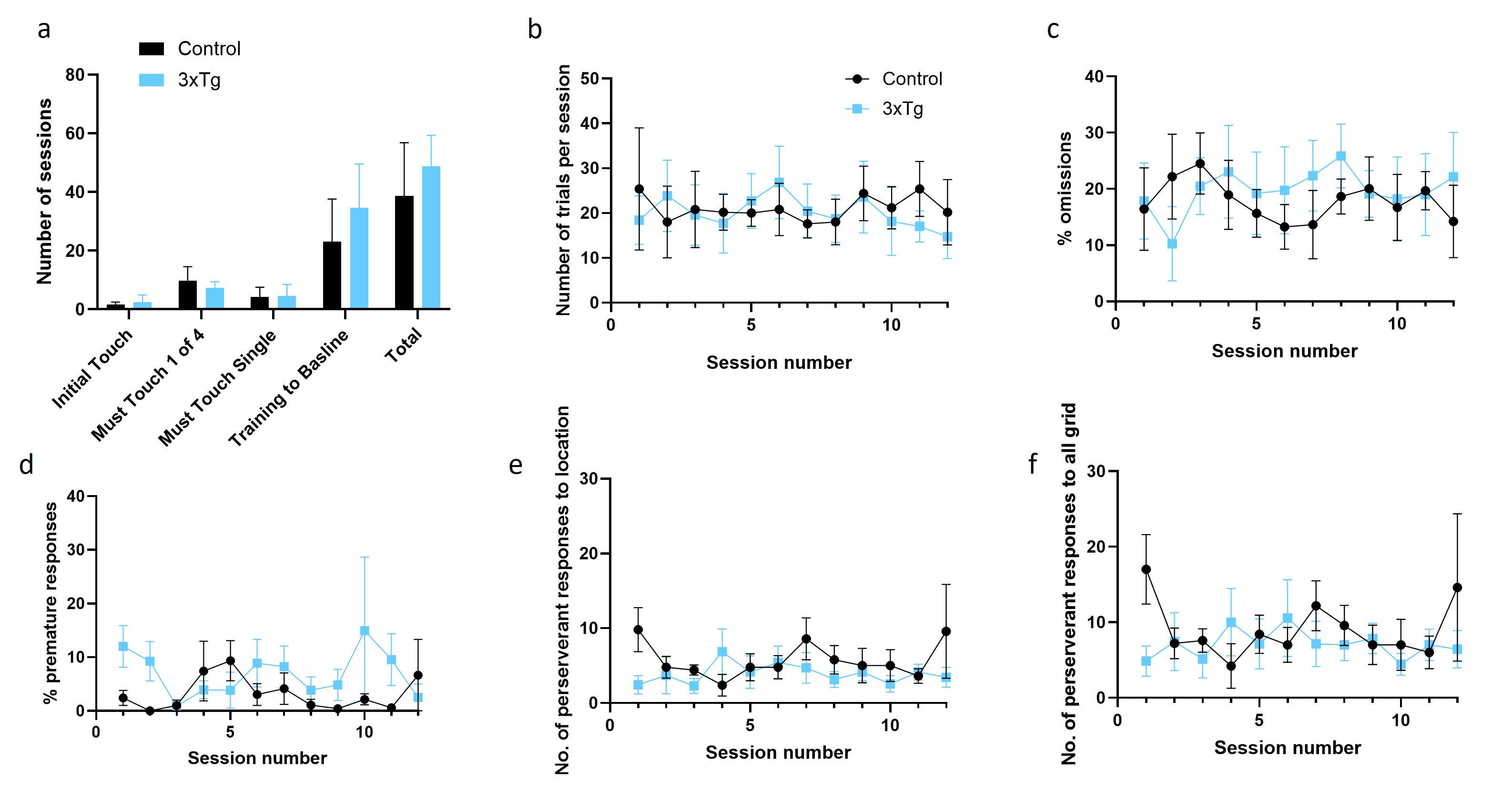
To assess executive function in an AD context, male 3xTg and control mice were trained to criterion on the 4-Choice Gambling Task (4CGT, Fig. 1), a rodent analogue of the Iowa Gambling Task (IGT) used to assess decision-making in the clinic. Mice began staged training at 2–3 months of age and were fully trained to criterion by 6–7 months of age (n = 5 controls, n = 7 3xTgs). The training performance of both groups was similar, with no significant difference in both the overall number of sessions required to reach criterion and the number of sessions required to reach criterion for each training stage. Similarly, during testing, the number of trials completed per session by 3xTgs was similar to controls. Additionally, the percentages of omissions, premature responses, and number of perseverant responses to the choice location or rest of the grid were similar between control and 3xTg mice. Therefore, 3xTg mice were able to learn and perform the task at a similar rate and engagement level to control mice (supplementary Fig. 1).
Analysis of test sessions revealed no significant effect of session in control (Fig. 2a) or 3xTg (Fig. 2d) mice, nor a significant interaction between session and average choice %. However, there was a significant effect of average choice % in control mice (Fig. 2a, F (3, 16) = 24.64; p < 0.0001) but not 3xTg mice (Fig. 2d). When sessions were combined (4 sessions per block), the significant effect of average choice % remained (Fig. 2b, F (3, 16) = 29.37; p < 0.0001). Control mice showed a clear preference for option P2, choosing it significantly more than other options in the first and second session blocks (Fig. 2b, p = 0.0424 for P2 over P3 in first session block; p < 0.0001 for P2 over P1 and P4, and p = 0.0003 for P2 over P3 in second session block). Conversely, 3xTg mice did not choose P2 significantly more than other options even following session blocking (Fig. 2e). When combining advantageous versus disadvantageous options (P1 + P2 versus P3 + P4), control mice showed a clear preference for advantageous options (Fig. 2c, F (1, 22) = 146.2; p < 0.0001) and chose these significantly more than disadvantageous options in all three session blocks (p = 0.0010, p < 0.0001, p = 0.0041) whilst 3xTgs did not show any preference (Fig. 2f).
Performance on the 4CGT can also be assessed by comparing the net advantageous choice of control and 3xTg mice (24). This was calculated by subtracting the average % of disadvantageous choices (P3 + P4) chosen over the course of the session from the % of advantageous choices (P1 + P2) chosen. Mixed-effects analysis showed a significant effect of genotype on net advantageous choice; control mice had a significantly higher net advantageous choice across individual sessions, averaging 50.93% across 12 sessions, compared to 10.61% for 3xTg mice (Fig. 2g, F (1, 10) = 8,203; p = 0.0168). When sessions were blocked, this significant effect of genotype remained (Fig. 2h, F (1, 10) = 9.419; p = 0.0119). Multiple comparison analysis showed that controls had a significantly higher net advantageous choice in individual session blocks 1 and 2 (Fig. 2h, p = 0.0220 and p = 0.0289, respectively). Control mice were, therefore, able to establish an advantageous strategy over the first few trials, and then maintain this strategy for subsequent sessions, whereas 3xTgs could not develop an advantageous strategy (Fig. 2g). As other aspects of task performance were similar between groups (supplementary Fig. 1), the inability of 6-8-month 3xTg mice to perform the 4CGT was most likely due to cognitive dysfunction and deficits in decision-making, as opposed to a lack of motivation, atypical locomotor activity, or an inability to learn the task.
Altered input/output and paired pulse responses in the prefrontal cortex of 3xTg mice following stimulation of hippocampal inputs.
To examine how vHIP-mPFC connectivity may be altered in 3xTg mice, local field responses were recorded in brain slices from layers II/III and V of the IL cortex following stimulation (10 to 100V intensities) of hippocampal fibres innervating these regions (52) (Fig. 3a). Representative responses for control and 3xTg mice are shown in Fig. 3b and 3d, respectively. In IL layer II/III, input-output curves showed a significant effect of genotype (p < 0.0001) and stimulation strength (p < 0.0001) and a significant interaction (Fig. 3c, F (9,150) = 3.658; p = 0.0004). Pairwise comparisons showed that local field responses in IL layer II/III were significantly lower in 3xTg than control mice between 40 and 100V (Fig. 3c, p = 0.0432, p = 0.0053 and p = 0.0007 for 40V, 50V and 60V stimulations respectively, p < 0.0001 for 70V to 100V stimulations). Additionally, population spikes were more frequently observed in layer II/III of control animals compared to 3xTgs (Fig. 3f, F (3,129) = 92.06; p < 0.0001). These observations support a reduced synaptic connectivity between the vHIP and mPFC, and dampened spread of postsynaptic excitation within the mPFC of 3xTgs. However, the presence and latency to maximal amplitude for a slower (presumed inhibitory) response component in layer II/III were similar between control and 3xTg LFPs (Figs. 3g and 3h). Response amplitudes in IL layer V were similar between 3xTgs and control mice (Fig. 3e).
Next, to further explore the synaptic short-term plasticity of hippocampal-prefrontal synapses, responses were recorded following paired pulse stimulation of innervating hippocampal fibres at 20, 50, 100, 200, and 500ms intervals. As with IO responses, paired pulse responses generally exhibited an initial excitatory local field response, followed by a slower presumed inhibitory component. In layer II/III, control mPFC LFPs exhibited mild paired pulse facilitation (PPF) at all intervals except 20ms, whereas 3xTg LFPs displayed a PPR very close to 1 (i.e., no facilitation) at all intervals except 20 ms, at which they exhibited paired pulse depression (PPD). However, 2-way ANOVA analyses of PPRs in layer II/III did not show a significant effect of genotype or interval, nor a significant interaction (Fig. 4a and 4b). Conversely, in layer V, a significant effect of genotype (F (1, 50) = 40.79; p < 0.0001) and a significant interaction between genotype and interval was observed (Figs. 4c and 4d, F (4, 50) = 3.756; p = 0.0095). Multiple comparisons analysis showed that PPRs at shorter intervals (20, 50 and 100ms) were significantly reduced in 3xTg compared to control mice (Fig. 4d, p < 0.0001, p = 0.0002 and p = 0.0498 respectively). At all five intervals, 3xTg responses exhibited PPD, indicative of a reduction in short-term synaptic plasticity in the vHIP-mPFC layer V input in 3xTg mice.
3xTgs displayed gene expression changes associated with glutamatergic, GABAergic, and dopaminergic activity, and calcium ion transport.
To determine whether the behavioural and electrophysiological changes described above were associated with any significantly enriched genes at similar preclinical stages, bulk RNA sequencing analyses of the mPFC was performed in 3- and 6-month-old 3xTg and control male mice. Genes were grouped using the gene ontology (GO) major bioinformatics classification system, which contains cellular component, molecular function, and biological process branches. In general, gene expression changes were not as numerous or large in 3-month compared to 6-month animals, and no descriptive terms were significantly enriched in 3-month mice; Fig. 5a displays the number of genes associated with each term (the ‘Count’), the ratio of the number of differential genes linked with the term to the total number of differential genes (the ‘Gene ratio’), and the significance between the two groups given by the adjusted p-value (padj). Individual gene expression changes associated with neuronal synaptic function and neurotransmitter release were of particular interest in this study, as these are likely to underlie alterations to IO response amplitudes and PPRs. Seven genes of interest associated with these terms were identified as differentially expressed genes (DEGs) between 3xTg and control mice at 3 months of age: Gpm6b, Slc38a5, Ccr5, Kcnj10, Ddah1, Gad1 and Slc17a8/VGLUT3. When plotted in a clustered heatmap, high levels of hierarchical clustering were observed between 3xTg and control mice for these genes (Fig. 5b). Several of these are associated with glutamatergic activity and transport, including Slc38a5 (SNAT5), Slc17a8 (VGLUT3), GAD1 and Kcnj10, whilst Gpm6b and Ddah1 have been linked with neurotransmitter activity and metabolism. Ccr5 is a chemokine receptor that has been reported to play a role in calcium ion transport and glutamatergic function. The fold change of each of these genes in 3xTgs compared to controls is shown in Fig. 5c. Interestingly, substantial differences in the expression of genes associated with cytosolic calcium transport were not observed in 3-month-old animals, even though associated genes showed a high level of differential expression by 6 months. Thus, mPFC gene changes underlying attenuated glutamatergic synaptic transmission occur earlier in disease presentation than changes to expression of genes associated with calcium ion transport.
More substantial gene expression changes were observed between 6-month-old control and 3xTg animals. Descriptive terms with most significant gene changes were related to immune function and defence responses, specifically the production of interleukins-1 and 6, tumour necrosis factor and MAPK and ERK1/2 kinase activity (Fig. 6a). Significant gene changes were also observed in several descriptive terms linked with AD, such as ‘response to lipoprotein particle’, ‘divalent metal ion transport’, and ‘negative regulation of neurogenesis’ (Fig. 6b). When terms associated with neuronal synaptic function and neurotransmitter release were compiled (Fig. 6c) and gene expression between controls and 3xTgs compared, terms associated with glutamatergic and GABAergic synaptic transmission, neurotransmitter function and transport, and calcium ion transport were significantly enriched (Fig. 6d). Genes associated with these descriptive terms were plotted on a clustered heatmap (Fig. 6e), which revealed high levels of hierarchical clustering between 3xTg and control samples, indicating large variability between control and 3xTg mice in the gene expression present within these descriptive terms.
Next, specific gene expression changes associated with the above significant terms in 6-month animals were investigated in more detail. Terms relating to glutamatergic function, including ‘glutamate binding’ (padj = 0.0222), ‘glutamatergic synaptic transmission’ (padj = 0.0175) and ‘negative regulation of glutamatergic synaptic transmission’ (padj = 0.0113) were significantly enriched between groups. In 3xTgs, genes encoding for glutamatergic receptor subunits, such as Grin2d, Grik1, Grin2a and Gria4 were downregulated, as was Cps1 associated with glutamate binding (Fig. 7a). Genes associated with glutamatergic synaptic transmission, such as Slc17a8 (encoding the vesicular glutamate transport 3; VGLUT3), Prkn and Unc13c were significantly downregulated, whilst Adora2a, Plat, Drd1 and Drd2 were upregulated (Fig. 7b). Genes associated with negative regulation of glutamatergic synaptic transmission, such as Npy2r, Adra1d, Htr2a and Grik1/2, were downregulated (Fig. 7c).
Changes to gene expression associated with GABAergic transmission were also plotted, to establish how changes to GABAergic function underlie alterations to the E-I balance. ‘GABAergic transmission’ was a significantly enriched term (padj = 0.0165), and expression changes were observed in multiple genes including Erbb4, Phf24, Nlgn1 (downregulated), Rac1 (upregulated) and receptor subunits including Gabra1, Gabrb2, Gabrg2, Gabrg3, Adra1a, Adora2a and Drd2, most of which were downregulated in 3xTgs (Fig. 7d). Due to the frequent observation of gene expression changes to neurotransmitter receptor subtypes, further terms associated with neurotransmitter receptor function were also investigated, and ‘dopamine transport’ (padj = 0.0265) ‘neurotransmitter receptor activity’ (padj = 0.0296), and ‘neurotransmitter metabolic process’ (padj = 0.00367) were all significantly enriched descriptive terms in 6-month-old 3xTg mice (Fig. 7g-i). Significant gene expression changes associated with these terms included the downregulation of Prkn and upregulation of Drd1 and Drd2 in 3xTgs.
Finally, to identify underlying gene changes associated with increased vesicle release probability in 3xTgs, as was observed in paired pulse stimulation experiments, gene expression changes underlying calcium ion transport were plotted. ‘Calcium ion transport’ (padj = 0.00284) and, more specifically, ‘cytosolic calcium ion transport’ (padj = 0.0152) were significantly enriched terms. Individual gene changes revealed a general, strong upregulation of genes involved in regulating calcium ion transport in both neurons and glia, including Ccl5, Cxcl10, Lyn, Icam1, Gem and Ifg1 (Fig. 7e, f). Descriptive terms linked with vesicle release machinery, including ‘presynaptic membrane organisation’, ‘presynaptic membrane’, ‘synaptic vesicle fusion to presynaptic active zone membrane’ and ‘presynaptic process involved in chemical synaptic transmission’ were not significantly enriched. This suggests that observed alterations to the vesicle release probability in 3xTgs are due to changes in calcium ion transport in the presynaptic membrane, as opposed to variable expression of proteins associated with vesicle release.
{kind=link}