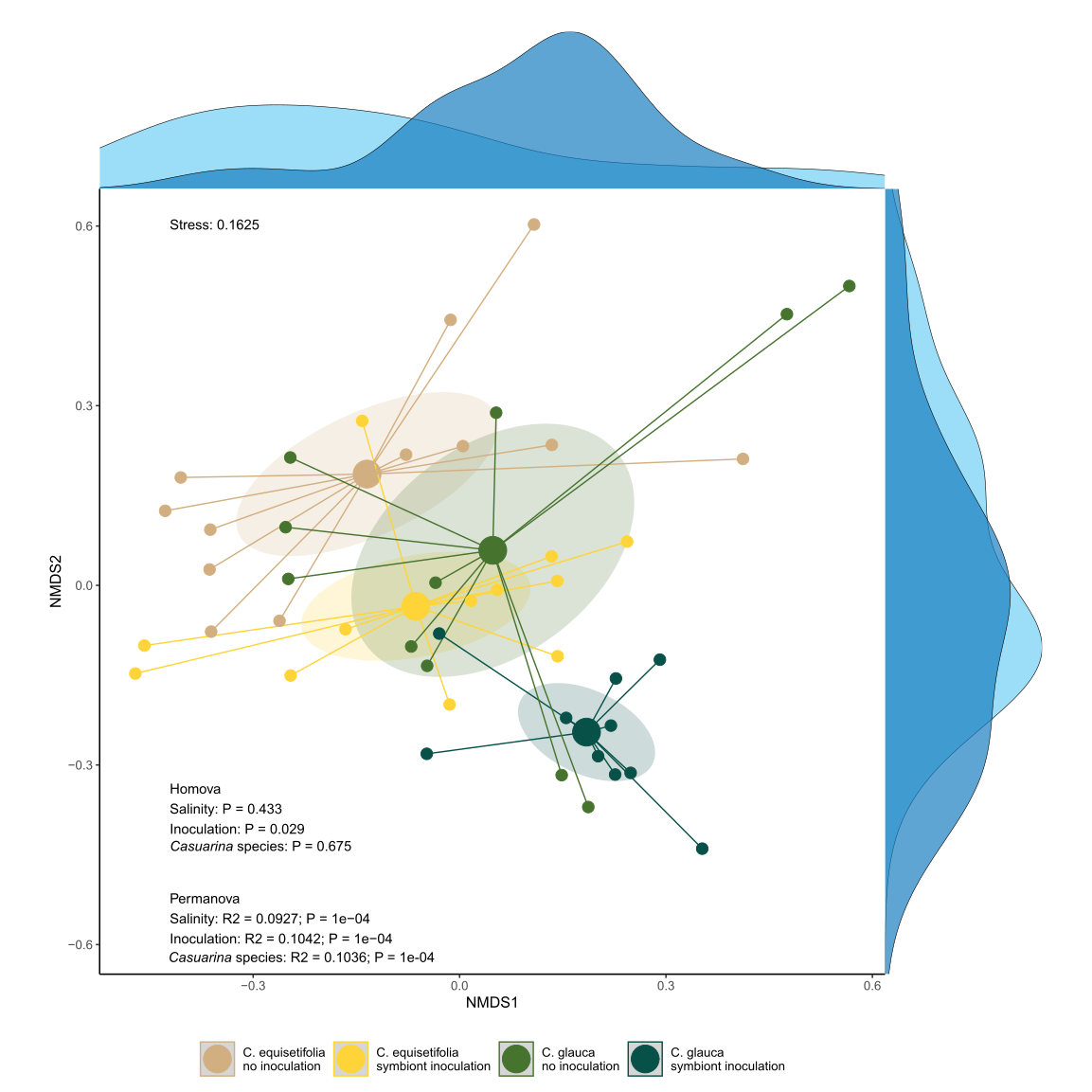
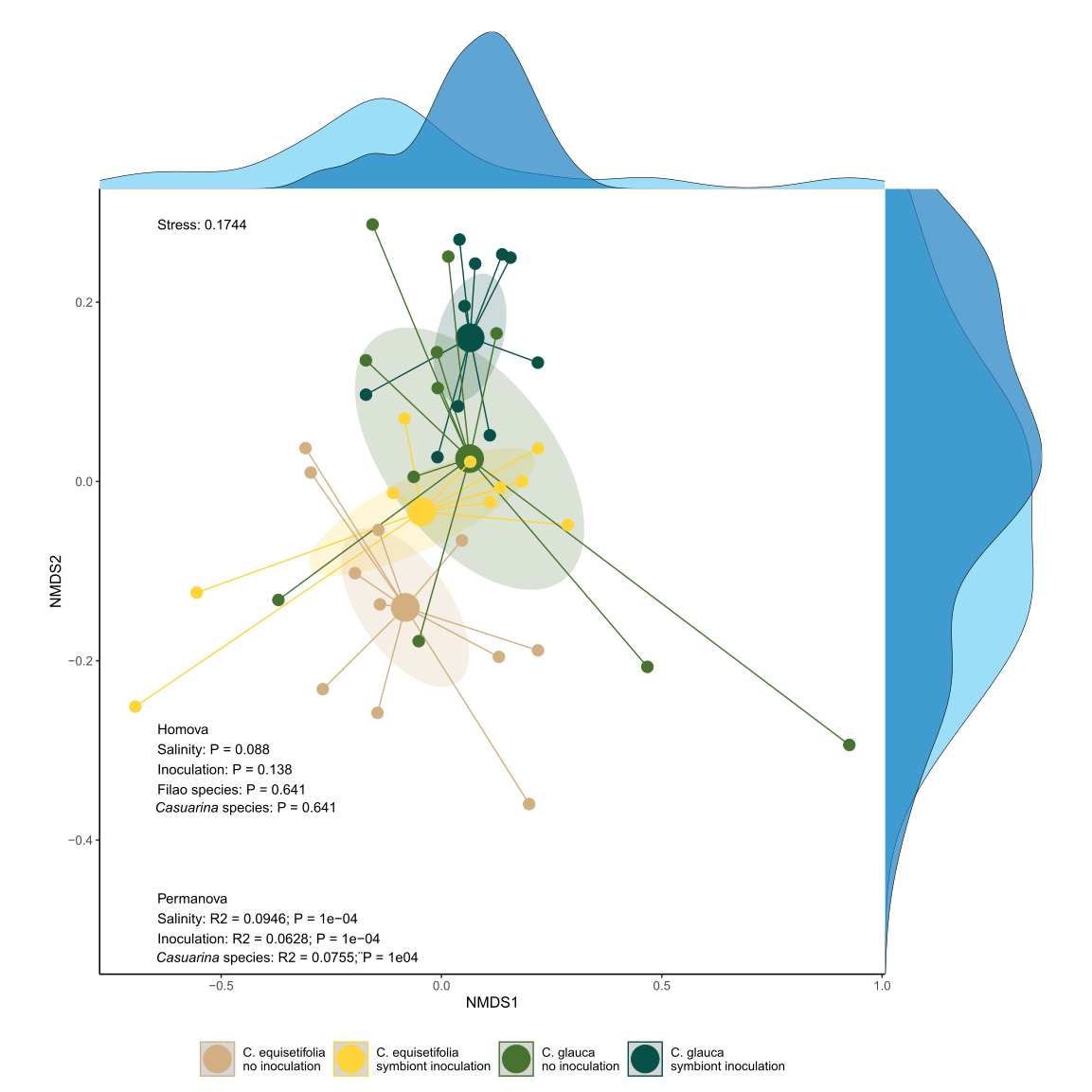
Salinity is expected to be one of the main factors affecting ecosystem functioning, notably soil microbiota (Lozupone and Knight, 2007; Rath et al., 2019; Yang and Sun, 2020; Zhao et al., 2019). Consequently, ecological strategies based on the use of bioinoculants are developped to counteract negative effects of salt on plant growth and survival. Yet adverse impacts of bioinoculants on soil functioning were observed (Cornell et al., 2021; Gou et al., 2024), with in most cases no resilience of the native microbial community (Mawarda et al., 2020). In the current study, salinity was a major factor of bacterial diversity changes, but symbiont inoculation was the main factor for fungal diversity.
Salinity was described as a key environmental factor driving bacterial community diversity in arid ecosystems (Zhang et al., 2019), but differential impacts are observed depending on salinity ranges (Zhang et al., 2019). Similarly, salinity and pH were shown as strongly correlated to bacterial community diversity, but inversely for pH compared to previous studies (Zhao et al., 2018). Differences in pH gradient might explain these opposite effects, since acid pH gradient (3 < pH < 6) was measured in the current study compared to alkaline pH gradient (7.9 < pH < 8.7) in Zhao et al. (2018). The difference in response between bacteria and fungi along a salinity gradient has been also described in other arid ecosystems (Lin et al., 2023), and contrasting toxicity effects of salt were between bacteria and fungi (Rath et al., 2016). The lack of effect of salinity gradient on the diversity of the fungal community might also be due to other environmental factors outweighing the effect of salinity (Zhao et al., 2019). The presence of vegetation cover, for example, could be one of those since the presence of cover plants can mitigate the effect of salinity on soil microbiota (Dasgupta et al., 2023). The plantation of Casuarinaceae on the current rehabilitation site was indeed shown to enhance the development of herbaceous vegetation (Djighaly et al., 2020). In addition, soil fungal diversity tend to be more closely aligned with vegetation cover both for mutualists (Cassman et al., 2016; Zhang et al., 2021) and saprobes (Francioli et al., 2020), thus less changes are expected in their communities without concurrent changes to vegetation (van der Heijden et al., 1998; Beck et al., 2020).
On the other hand, changes in diversity of the predominant bacterial taxa have been observed between Casuarina species, which might be due to differences in root traits (exudates, genetic, tolerance to stress) between the two species. Bacterial community have been indeed shown as more strongly affected by differences in root traits than fungi (Alahmad et al., 2024; Merino-Martín et al., 2020). The changes in vegetation cover (richness diversity, composition) due to the different environmental factors (salinity, symbiont inoculation, Casuarina species) remains unknown until now, but constitute undoubtedly an important explaining factors to explain variations in soil microbiota (Djighaly et al., 2020). However, the composition of each microbial compartment was globally influenced by the various environmental factors, suggesting that the use of bioinoculant, as well as the type of Casuarina species determine the characteristics of community assembly in addition to salt stress. The consequences in terms of microbial functionalities and ecosystem services could be important (Catano et al., 2023), and have to be taken into account for rehabilitation programs.
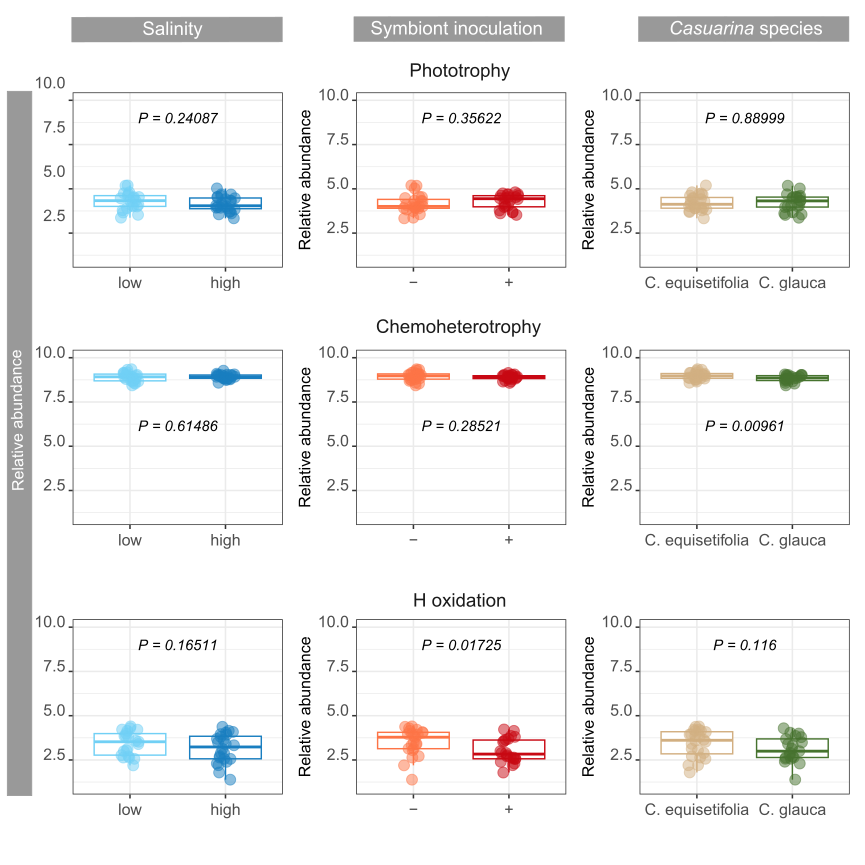
Bacteria and fungi play indeed major roles in soil biogeochemical cycles, supporting soil fertility and plant health (Banerjee and van der Heijden, 2022; Saleem et al., 2019; Trivedi et al., 2020). Modifications of biotic and abiotic parameters can have a serious and lasting impact on soil functioning by altering general and specialized microbial functional groups (Griffiths and Philippot, 2013; Xun et al., 2021). For example, salinity, pH, soil moisture, or nutrients were shown to contribute to changes in fungal trophic guilds in saline soils (Zhao et al., 2019). Current results revealed significant impacts on several microbial functional groups or guilds dependent on single or dual environmental factors.
Saprotrophs, symbiotrophs and pathotrophs were mostly impacted by symbiont inoculation and / or Casuarina species. Nevertheless, salinity affected the abundance of fungal plant pathogens, but not bacterial pathogens. Salt stress has also been shown to increase aggressiveness of certain fungal pathogens (Haddoudi et al., 2021). Opposite effects were observed on the abundance of actinorhizal N-fixers and free N-fixers in soils, highlighting complex plant-soil feedbacks, potentially due to priority effects between the competitive actinorhizal symbiont pre-inoculated on Casuarina seedlings and native actinorhizal community on the rehabilitation site, and plant-mediated promoting effect on other N-fixers. Nevertheless, N fixation in soils was reduced for high levels of salinity.
The use of AM fungal inoculant combined with actinorhizal inoculant could also impact the soil microbial community through plant-soil feedbacks by enhancement of host plant growth and modification of the amount, timing, and form of carbon inputs into soil (De Gruyter et al., 2022). AM fungal inoculation was hypothesized to modify the native AM fungal community, as frequently observed (Islam et al., 2021; Thioye et al., 2021, 2019), but the extent of changes are related to properties of AM fungal inoculant and environmental factors (Hart et al., 2017; Islam et al., 2021; Martignoni et al., 2020). In the current study, Casuarina species was the explicative factor of changes, with higher abundance of soil AM fungi associated with C. glauca compared to C. equisetifolia. Similar observation was obtained for native soil rhizobia. These results emphasize the necessity to set up trial with mixed plantation to evaluate the benefits of a range of Casuarina species on soil functioning beyond the effects on tree growth.
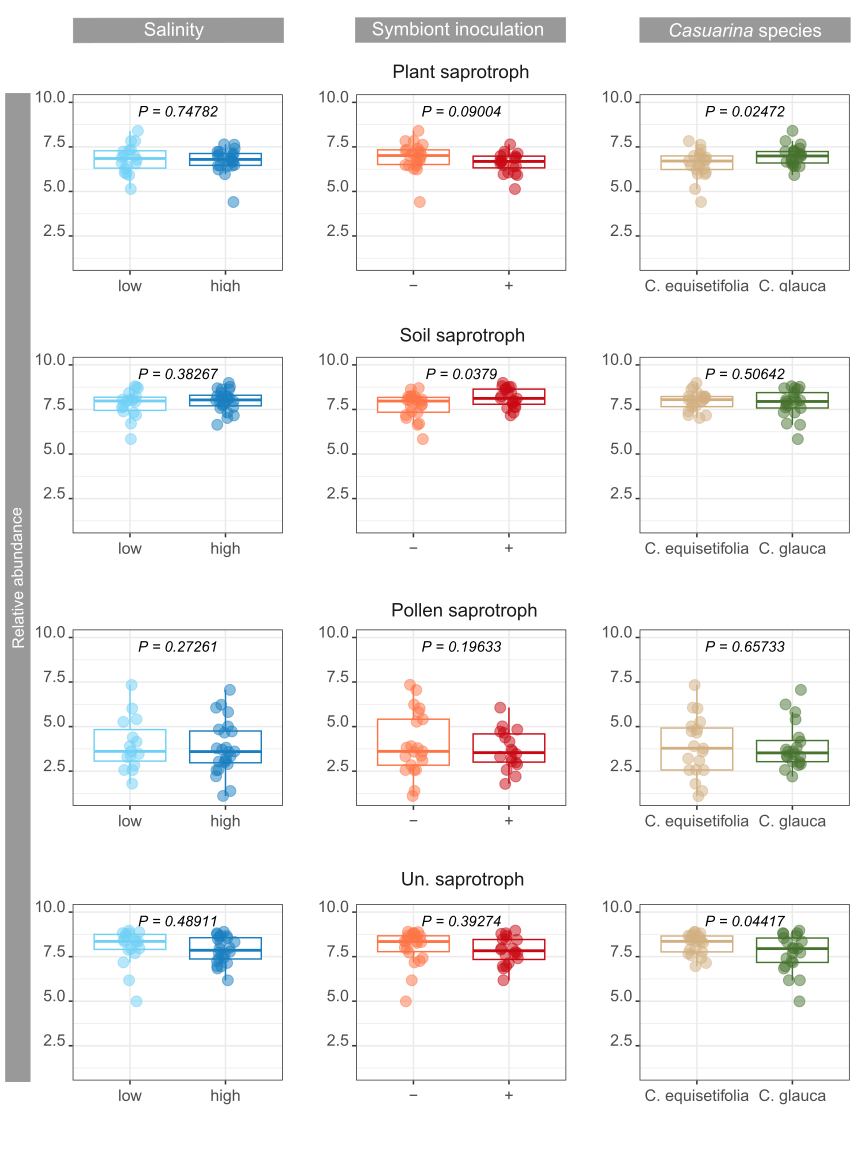
Changes in saprotroph abundance might be due to differences in vegetation cover promoted by Casuarina plantation and consequently differences in organic matter composition or abundance. In addition, differential impacts of Casuarina species were observed on bacterial and fungal saprotrophs. Symbiont inoculation impacted also differentially bacterial and fungal saprotrophs, favoring soil fungal saprotrophs and limiting plant bacterial saprotrophs.
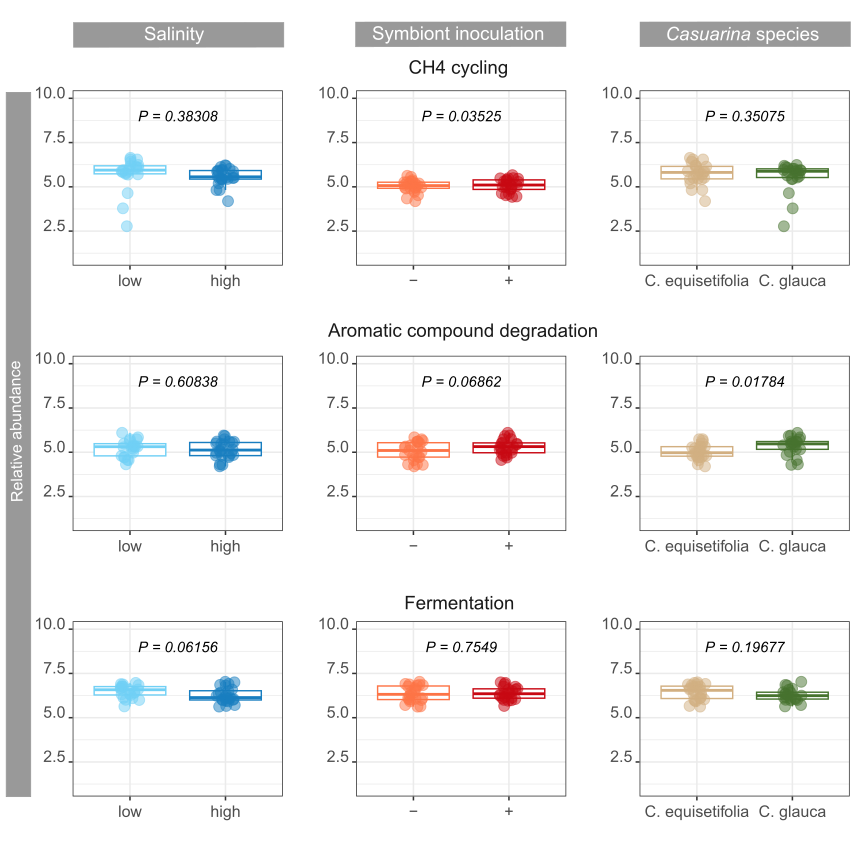
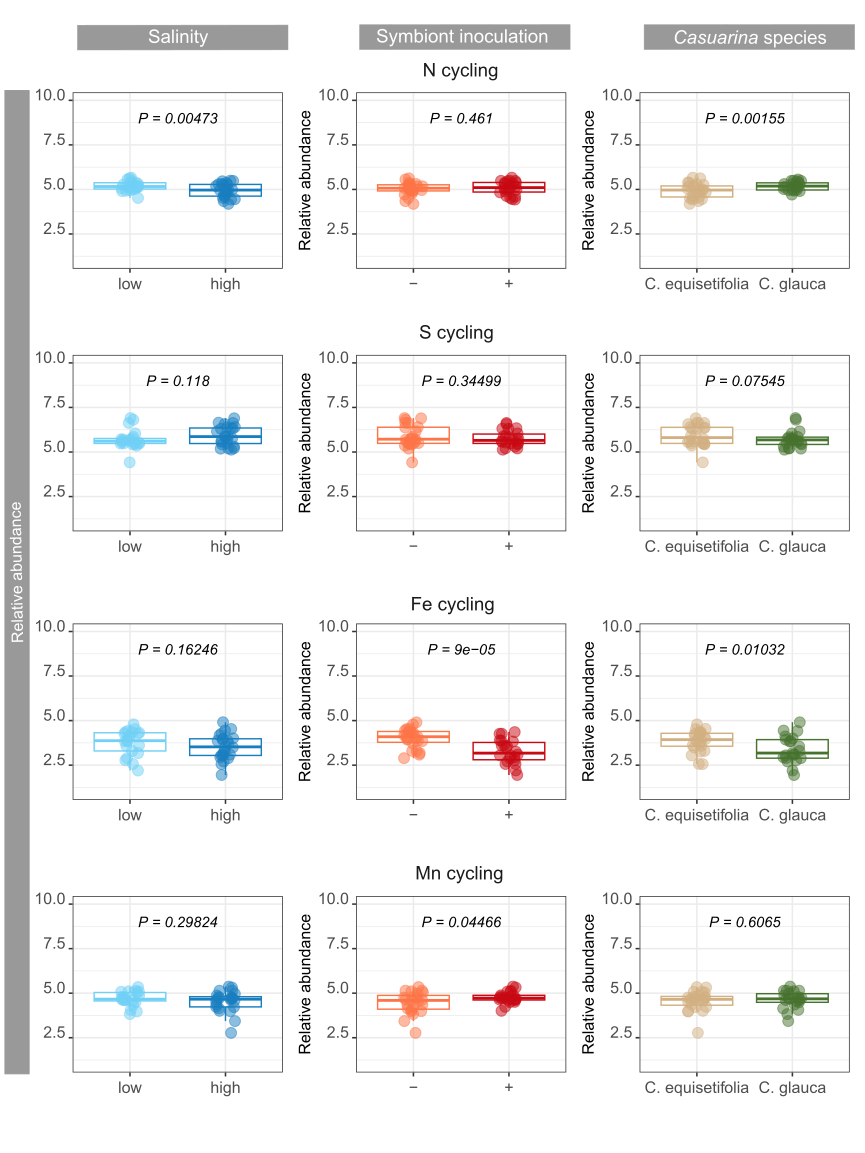
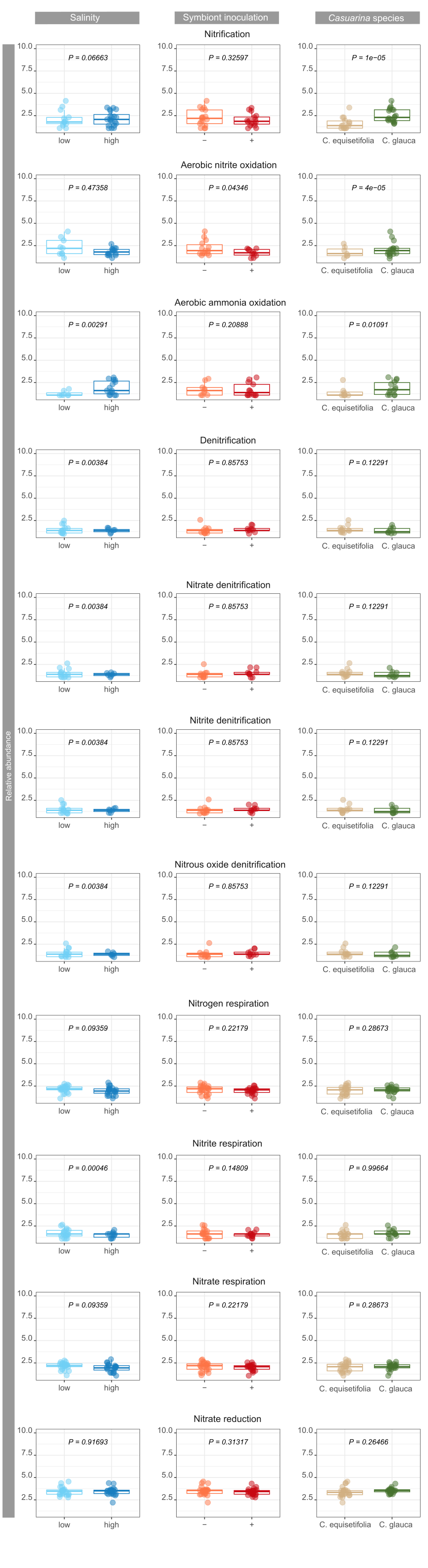
By digging deeper into the functions involved in biogeochemical cycles, significant impacts of environmental factors were observed on nutrient cycling and energy flows. Soil carbon content, notably CH4 cycling and aromatic compound degradation, was affected by symbiont inoculant and Casuarina species, respectively. Multiple microbial guilds are responsible of CH4 fluxes in soils (Hartman et al., 2024), and the strong changes of soil bacterial community due to symbiont inoculation might have particularly impacted some of the guilds involved in CH4 cycling. Differences in the abundance of bacteria involved in aromatic compound degradation is probably due to differences in root exudate composition and rates between the two Casuarina o species (Dennis et al., 2010; Staszel-Szlachta et al., 2024). On the other hand, salinity was the main factor negatively impacting soil bacterial community mediating N cycling confirming previous studies in highly salt-affected soils (Li et al., 2021). Nevertheless, impacts on N turnover is dependent on salinity levels, which can lead to inconsistency in conclusions (Tao et al., 2024). Moreover, changes in H-oxidizing bacteria abundance, which are generally associated with N fixation variations (Maimaiti et al., 2007), was not related to salinity levels, but to symbiont inoculation with opposite effects on N fixation. Bioavailability of soil N content for plant was also promoted by symbiont inoculation through ureolytic bacterial community, with probable major benefits on soil fertility (Hasan, 2000). Interestingly, the use of C. glauca promoted bacterial community involved in N cycling compared to C. equisetifolia, from N fixation to nitrification, whereas increasing salinity promoted mainly denitrification.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}