1. F. nucleatum promoted periodontal and intestinal destruction in mice.
UC model was constructed by free-drinking 2.5%DSS solution and the periodontitis model was constructed by F. nucleatum periodontal infection (Fig. 1A). Compared to gCON and gFN, the survival rates of mice in gUC and gUF declined to 90% and 70%, respectively (Fig. 1B). The weight of mice in gCON showed a continuous increase, fluctuated around the initial weight in gFN, decreased significantly in gUC, and decreased further in gUF (Fig. 1C). The disease activity index (DAI) score elevated significantly in gUC and gUF compared with gCON and gFN, and was the highest in gUF (Fig. 1D). In addition, colon length decreased progressively and statistically in four groups (Fig. 1E, 1F). The colon was then stained using hematoxylin and eosin (H&E) to assess its histopathological characteristics (Fig. 1G). In gCON, the colonic epithelium and crypts were intact and continuous. In gFN, the inflammatory cell infiltration was fragmented. In gUC, the crypt structure was disrupted and there was significant infiltration of inflammatory cells in the submucosal layer, with some epithelial damage. In gUF, a large number of crypts were missing, and inflammatory cell infiltration and epithelial disruption were more pronounced. Meanwhile, H&E scores showed an increasing trend between the four groups (p < 0.05).
Mice exhibited periodontitis symptoms after F. nucleatum periodontal infection. H&E staining showed scattered inflammatory cell infiltration in gUC compared to gCON, disruption of gingival epithelial integrity, disorganization of periodontal ligament arrangement, inflammatory cell infiltration, and alveolar bone resorption in gFN. In contrast, periodontal inflammation was most significant in gUF (Fig. 1H). Micro CT showed alveolar bone resorption in gFN, as evidenced by an increase in the cemento-enamel junction-alveolar bone crest (CEJ-ABC) distance and trabecular separation (Td.Sp.) and a decrease in bone tissue volume (BV/TV) and bone surface area-tissue volume ratio (BS/TV) in the mandibular first molar (p < 0.01). Mice in gUF exhibited the highest alveolar bone resorption (pCON−UF<0.001). Slight alveolar bone resorption was also observed in gUC, but it was not statistically significant (pCON−UC>0.05) (Fig. 1I, 1J).
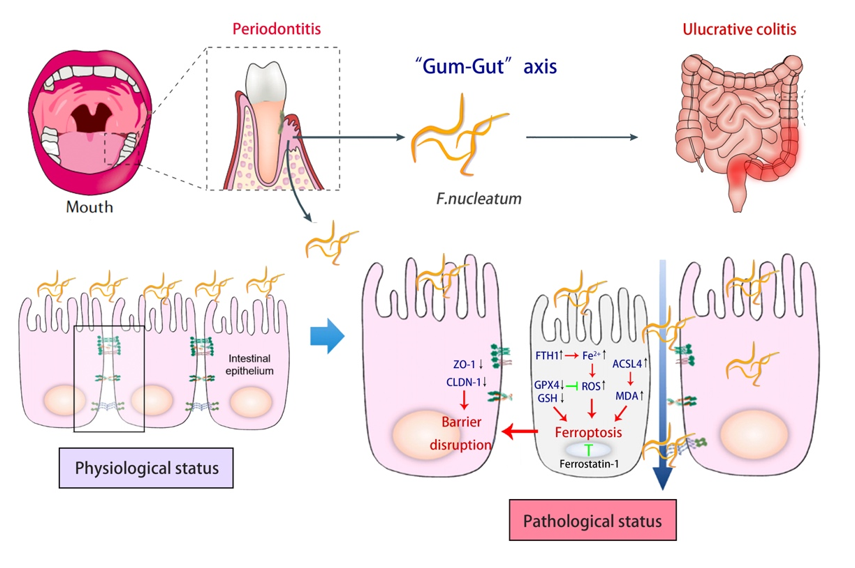
These data suggest that F. nucleatum is capable of destroying both periodontal and intestinal tissues, and may be a potential bridge symbiont for the association between periodontitis and UC.
2. F. nucleatum promoted intestinal barrier disruption in UC mice.
To confirm the invasive ability of F. nucleatum in the intestinal tract, we performed FISH experiment (Fig. 2A). The results showed that F. nucleatum from periodontal infections accumulated in the intestinal lumen and adhere to the IECs. In gFN, the number of invaded F. nucleatum in the lamina propria was limited (pCON−FN>0.05). In contrast, a large amount of F. nucleatum could be seen invading the deeper layers in gUF (pCON−UF<0.001). This suggests that the intestinal barrier is effective in isolating pathogenic F. nucleatum invasion in physiological states. Furthermore, F. nucleatum abundance was significantly higher in gUC intestinal tissue (pCON−UC<0.001). We also confirmed that elevated F. nucleatum levels in the colon of the gUF were oral in origin (Fig. S1 and Table. S3).
Next, we tested the intestinal barrier function. Alcian blue/periodic acid-Schiff, (AB-PAS) staining results showed that F. nucleatum aggravated intestinal mucus layer destruction in UC (pUC−UF < 0.05) (Fig. 2B). Transmission electron microscopy (TEM) revealed that the tight junctions (TJs) between the IECs were intact, and the intestinal villi were neatly arranged and moderately dense in gCON. Although the TJs appeared intact, there were fewer intestinal villi in gFN. In gUC, typical disruption of TJs and loss of villi were observed. While in gUF, the intestinal villi were largely absent, TJs were disrupted, the intercellular gap widened, and the organelles were detached (Fig. 2C). Serum FD4 concentration, representing intestinal permeability, increased substantially following F. nucleatum challenges (pUC−UF < 0.0001) (Fig. 2D). We then detected the expression of the TJs proteins Zona occludens 1 (ZO-1) and Claudin 1 (CLDN-1) using immunohistochemical staining (IHC) and western blot (WB). In comparison to gCON, ZO-1 and CLDN-1 protein expressions were lower in gUC and gUF (Fig. 2E, 2F), as well as mRNA (p < 0.05) (Fig. 2G).
These results showed that F. nucleatum could aggravate the intestinal barrier disruption in UC mice.
3. F. nucleatum promoted UC development associated with ferroptosis.
The colonic content of the mice was analyzed using 16S rRNA sequencing and LC-MS technique. The rarefaction curves flattened, and the sequencing data reached saturation, enabling coverage of most of the species. Gut microbial diversity was significantly reduced in gUF (Fig. S2). Beta diversity showed a clear trend of dispersion between gCON and gUF (Fig. 3A), as did the community abundance composition (Fig. 3B). Analysis of the species with significant differences based on LEfSe revealed that Fusobacteriaceae was one of the dominant bacteria in the gFN, gUC, and gUF (Fig. 3C). The sample-species relationships are shown in Fig. S3. The PICRRUSt2 functional prediction heat map showed that “cell growth and death” from level 1 “cellular process” was implicated in the differential pathway between four groups, with “necroptosis, ferroptosis, and apoptosis” as the specific mechanism (Fig. 3D).
Based on the metabolomic analysis, the PCA score plot revealed differences between gCON and gUC (Fig. 3E). The Venn diagram showed the different metabolite compositions between groups (Fig. 3F). Diagram after comparing and statistically mapping intestinal metabolites with categorization information from the HMDB 4.0 database was shown in Fig. 3G. The KEGG functional pathway identified “ferroptosis” and "apoptosis” as the main pathways involved in “cell growth and death” (Fig. 2H). KEGG pathway enrichment and iPath metabolic pathway analyses are shown in Fig. S4 and S5.
The combined 16S rRNA sequencing and LC-MS results suggested that the aggravation of UC by F. nucleatum infection may be associated with ferroptosis.
4. F. nucleatum induced ferroptosis in the colon of UC mice.
To confirm this hypothesis, we examined the expression of the ferroptosis regulators GPX4, FTH1, and ACSL4. IHC and WB of mouse colon showed decreased GPX4 protein levels but increased FTH1 and ACSL4 protein levels (Figs. 4A, 4B). RT-qPCR identified changes in the mRNA levels of GPX4, FTH1, and ACSL4, which were consistent with the protein results (Fig. 4C). Moreover, MDA and iron contents, particularly ferroptosis-related Fe2+, were upregulated in the colon, whereas the reduction ratio of GSH was downregulated (Fig. 4D-F). These results suggested that F. nucleatum infection increased susceptibility to ferroptosis and exacerbated colonic ferroptosis in UC mice.
Immunofluorescence staining revealed significantly altered positive signaling cells, primarily in the epithelium. We therefore performed double immunofluorescence staining for the epithelial cell marker CK18 and ferroptosis marker GPX4. The results revealed that the overlapping signals of CK18 and GPX4 were significantly reduced in gUC and gUF compared with gCON and gFN (Fig. 4G). Based on the above data, we hypothesized that F. nucleatum adheres to and induces ferroptosis in IECs, thereby disrupting the gut barrier.
5. F. nucleatum induced ferroptosis in the colonic epithelial cells.
To further clarify the effect of F. nucleatum on the IECs, we investigated ferroptosis in the normal colonic epithelial cell CCD841 (ATCC CRL-1790). DSS solutions and F. nucleatum suspensions were selected at appropriate concentrations and co-cultured with the cells for 24 h (Fig. S6). Cell viability and LDH release were then assessed. F. nucleatum significantly inhibited cell proliferation and caused cell damage (Figs. 5A, 5B). Furthermore, F. nucleatum promoted cell ferroptosis, as evidenced by increased MDA levels, decreased GSH ratios, and intercellular Fe2+ aggregation (Figs. 5C, 5D, and 5G). RT-qPCR and WB results showed decreased GPX4 and increased FTH1 and ACSL4 mRNA and protein levels, consistent with in vivo experiments. (Figs. 5E, 5F). The IHC staining results further confirmed the trend changes in GPX4, FTH1, and ACSL4 (Fig. 5H).
Next, we introduced the ferroptosis inhibitors Fer-1 and deferoxamine (DFO), determined their optimal concentrations (data not shown), and found that Fer-1 outperformed DFO in rescuing CCD-841 cell activity (Fig. S7). Fer-1 significantly inhibited F. nucleatum-induced intercellular ROS aggregation and mitochondrial membrane potential (MMP) decrease, which are characteristic of ferroptosis (Fig. 5I, 5J).
These studies suggested that F.nucleatum increased cell ferroptosis in IECs and that Fer-1 could partially rescue this process.
6. Ferroptosis inhibitor rescued F.nucleatum-induced intestinal barrier disruption in UC mice.
To validate the effect of inhibiting ferroptosis on the progression of UC with F.nucleatum infection, we added Fer-1 treatment to the original grouping, and the mice were modeled as shown in Fig. 6A. The safety of Fer-1 intraperitoneal injection has been tested (Fig. S8). Compared to gUC and gUF, the survival rates of mice in gUC + Fer-1 and gUF + Fer-1 increased from 85.7–100% and from 62.5–85.7%, respectively (Fig. 6B). From day 25 of modeling, the weight of mice in gUC + Fer-1 and gUF + Fer-1 were significantly higher than those in gUC and gUF (p < 0.05) (Fig. 6C). The DAI score was significantly decreased in gUF + Fer-1 compared to gUF and even lower than that of gUC + Fer-1 after 31 days of modeling (p < 0.001) (Fig. 6D). Colon length was significantly increased in gUC + Fer-1 and gUF + Fer-1 compared to gUC and gUF (p < 0.01). In addition, there was no statistically significant difference between gUC + Fer-1 and gUF + Fer-1 (Fig. 6E, 6F). These results suggested that inhibition of ferroptosis is effective in suppressing disease progression in UC with F.nucleatum infection.
Next, we measured the intestinal barrier following the inhibition of ferroptosis. Serum FD4 concentration was significantly lower in gUF + Fer-1 compared with gUF (p < 0.0001) but was not statistically different in gUC and gUC + Fer-1 (Fig. 6G). The histopathological changes were evaluated using H&E staining and AB-PAS staining, as shown in Fig. 6H, and 6I. Compared to gUC and gUF, lighter epithelial damage, fewer inflammatory cell infiltration, and less crypt disruption as well as increased mucus production were observed in gUC + Fer-1 and gUF + Fer-1. WB and IHC staining showed that ZO-1 and CLDN-1 protein expressions were significantly upregulated in gUC + Fer-1 and gUF + Fer-1 compared with gUC and gUF (p < 0.05) (Fig. 6J, 6K). These results proved that the application of ferroptosis inhibitors can significantly rescue intestinal barrier disruption aggravated by F.nucleatum.
{kind=link}