Inhibition of macrophage motility reduces Py8119 mammary tumor growth
To determine whether HCK activity regulates macrophage motility and associated invasive growth, we took advantage of the fact that HCK becomes activated in response to ligand engagement of the CSF-1R-and resulting stimulation of macrophage motility. Accordingly, we inhibited HCK activity with the pyrrolo-pyrimidine compound RK20449 (25), which we have previously shown to block motility and invasive capacity of both WT and HckCA BMM in vitro (10, 13), and to reduce tumor growth in colorectal, gastric and pancreatic cancer in vivo (13–15). Female mice were injected with Py8119 cells and RK20449 injections started immediately, thereby reducing mammary tumor growth by 70% (3.3-fold) (Fig. 1A). We then observed that inhibition of PI3K p110δ with the selective inhibitor GS-9820 (acalisib) reduced tumor size by 35% (1.5-fold) (Fig. 1B) (26).
Figure 1. HCK and PI3K p110δ regulate Py8119 mammary tumor growth. (A) Tumor weights of mice treated prophylactically with the HCK inhibitor RK20449 (RK). (B) Tumor weights of mice treated prophylactically with the PI3K p110δ inhibitor GS9820 (GS) or the CSF-1R inhibitor GW2580 (GW). (C) Tumor weights of mice treated therapeutically with the HCK inhibitor RK20449 (RK). (D) Tumor sections stained for IBA1 + TAMs. (E) Quantification of TAMs at the tumor margin in vehicle and RK20449-treated tumors and at the tumor margin and tumor core in vehicle and GS9820-treated tumors. (F) Tumor weights of HckWT and HckKO mice. (G) Tumor weights of HckWT and HckCA mice. Scale bar, 200µm. Mean ± SEM is shown, ns denotes not significant, *p < 0.05, ****p < 0.0001.
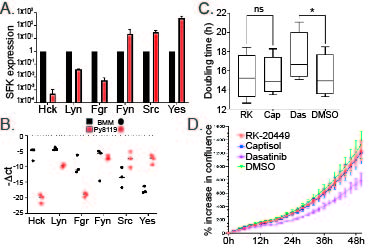
In contrast and consistent with previous reports, pan-CSF-1R inhibition by GW-2580 did not reduce tumor growth (Fig. 1B) (27). Finally, therapeutic inhibition of HCK by administration of RK20449 to mice with established (palpable) tumors also reduced tumor growth by 70% (Fig. 1C). To exclude the possibility that RK20449 treatment would directly affect the growth of Py8119 cells, which do not express HCK, we determined their doubling time and found that it was unaffected by RK20449 but suppressed by a pan-SFK inhibitor dasatinib (Supplementary Fig. 1A-D).
Supplementary Fig. 1. Py8119 cell in vitro data. (A,B) Relative expression of SFKs in Py8119 cells. (C,D) Proliferation of Py8119 cells treated with RK20449 or dasatinib, *p < 0.05.
To determine the effect of motility inhibition on TAM numbers in Py8119 tumors, we used immunohistochemistry (IHC) and the macrophage marker IBA1 to monitor the distribution of TAMs across Py8119 tumors. Abundant TAMs were seen, particularly at the tumor edge. While PI3K p110δ inhibition reduced the number of TAMs at the tumor edge, HCK inhibition neither affected their numbers nor their accumulation at the tumor edge (Fig. 1D,E). Taken together, inhibition of CSF-1R-activated macrophage motility but not full CSF-1R blockade reduced Py8119 mammary tumor growth. Moreover, tumor growth inhibition occurred independent of the number of distribution of TAMs.
HCK activity correlates with Py8119 mammary tumor growth
Because HCK inhibition profoundly reduced Py8119 tumor growth and hyper-motile HckCA BMM showed increased invasion in Py8119 mammospheres in vitro (13), tumor growth was examined in hosts either deficient in HCK expression (HckKO) or expressing constitutively active HCK (HckCA). Hck-deficient hosts limited growth by more than 7-fold compared to WT hosts whereas hosts expressing the HCKCA isoform enabled tumors to grow to 4-fold increased size relative to WT hosts (Fig. 1F,G). Notably, frequent ulceration into overlying skin in HckCA mice necessitated sacrifice at smaller tumor size in this experiment.
Loss of HCK activity reduces SFK activity in Py8119 tumor margins
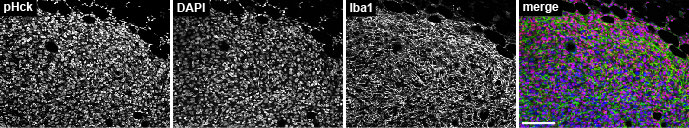
Because HCK activity correlated positively with tumor growth, we used a pY410-specific HCK antibody to examine the distribution of the active isoform of HCK by IHC. Strong nuclear staining in HckKO tumors indicated the antibody was not specific for HCK (Supplementary Fig. 2), whereas a pan-pY416-SFK specific antibody demonstrated plasma membrane-associated SFK activity at the margins of Py8119 tumors in treatment-naïve and vehicle-treated HckWT hosts (Fig. 2A-C). In contrast, membrane-associated SFK activity was absent at the margins of tumors recovered from HckKO and RK20449-treated HckWT hosts (Fig. 2A,C,D,F). Membrane-associated SFK activity extended further from the tumor margins in HckCA tumors although activity at tumor margins was not measurably increased (Fig. 2B,E). These results indicate that membrane-associated SFK activity is seen in TAMs and other cells at Py8119 mammary tumor margins, particularly in areas associated with invasion, and that much of the pY-SFK signal is due to active HCK in TAMs.
Supplementary Fig. 2. Anti-pHck antibody is non-specific. pY HCK (red) co-localizes with DAPI (blue) in Py8119 tumor nuclei in HckKO mice. Iba1+ TAMs are shown in green. Scale bar, 200µm.
HCK regulates cytotoxic T cell numbers in Py8119 tumors
We have shown in other tumor models that HCK deletion or inhibition increases cytotoxic T cell recruitment and activation (8, 15). This may account for some of the effect conferred by HCK expression in the host to the growth of Py8119 tumors. Consistent with this, both CD8+ T cell abundance and activity (perforin) were increased 4-fold in tumors from HckKO hosts and scant CD8+ T cells were seen in tumors from HckCA hosts (Fig. 3A-D). RK20449 did not change CD8+ T cell numbers (Fig. 3E), perhaps due to its known affinity for the T cell-specific SFK member LCK. Thus, HCK activity in TAMs appears to regulate cytotoxic T cell numbers and activity in Py8119 tumors.
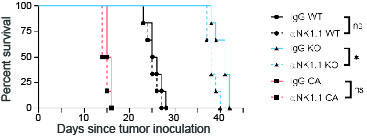
An anti-CD8+T cell antibody (YTS169) was used to examine the effect of CD8+ T cells on Py8119 tumor growth. HckWT, HckKO and HckCA hosts were injected with the antibody prior to, during and after tumor cell inoculation. CD8+ T cell depletion reduced survival of tumor-bearing HckWT hosts from 25.5 to 20.5 days (p < 0.01), from 41.0 to 34.0 days (p < 0.001) of HckKO hosts, and from 15.5 to 14.5 days (p < 0.05) of HckCA hosts (Fig. 3F). However, following T cell depletion, HckKO hosts survived 14 days longer than HckWT hosts lacking cytotoxic T cells (p < 0.0001) and 8.5 days longer than HckWT mice treated with an isotype IgG control (p < 0.001). NK cell depletion had little effect on tumor growth in any genotype (Supplementary Fig. 3). These results demonstrate that the effect of HCK inhibition is in part mediated by adaptive immune responses, which is consistent with our findings in MC38 colon and KPC pancreatic tumors (8, 15).
Supplementary Fig. 3. NK cell depletion does not affect Py8119 tumor growth. Survival of Py8119 tumor-bearing HckWT, HckKO and HckCA mice treated with either anti-NK.1 antibody or IgG isotype control.
TAMs comprise more than 40% of Py8119 mammary tumor mass
To further examine the role of HCK in Py8119 tumors, single cell RNA sequencing (scRNA-seq) was carried out on tumors harvested from RK20449-treated, HckKO, HckCA and matched control hosts. Initially, because the rapid growth and high ulceration rate of tumors in HckCA hosts resulted in the recovery of smaller tumors from matched WT hosts (0.206g) than those from WT hosts paired with HckKO hosts (1.04g), 10 day (early) and 14 day (late) tumors from WT hosts were characterized to determine whether tumor size affected tumor cellular composition. Transcriptomes of 16,477 cells were sequenced, filtered and normalized to cluster cells in an unbiased manner with annotation based on differentially expressed genes (DEGs) in Seurat and lineage specific markers (Supplementary File 1). TAMs comprised 42.6% of cells in early tumors and 42.1% in terminal volume tumors and tumor cells 43.5% and 39.1% respectively (Table 1) (Fig. 4A).
As tumor stage did not affect Py8119 tumor composition, tumors from all genotypes, treatment groups and stages were then combined (> 103,000 cells), clustered and annotated. Overall, TAMs made up 44.0% of cells compared to 44.4% tumor cells, 5.5% T and NK cells with small clusters of other cell types (Table 1). High Py8119 cell expression of two macrophage chemokines, CSF-1 and interleukin (IL)-34, led to TAM recruitment (Fig. 4B). When the scRNA-seq data were separated by HCK treatment or genotype, consistent with the IHC findings, HCK inhibition and deletion did not reduce TAM numbers (Table 1). In contrast, TAM numbers appeared to be reduced in HckCA tumors (Table 1). Also consistent with the IHC findings, HCK deletion doubled T and NK cell numbers (4.6 to 10.6%, p = 0.039) while RK290449 treatment reduced them (6.7 to 2.9%), the latter finding probably reflecting off-target inhibition of LCK in T cells. Finally, scRNA-seq data from tumors from HCK replete hosts confirmed that TAMs and DCs expressed HCK and other myeloid SFKs while T and NK cells expressed LCK and FYN (Fig. 4C). Py8119 cells expressed low levels of the ubiquitous SFKs only (Fig. 4C).
Py8119 tumor TAMs are pro-tumoral and cluster into five subtypes
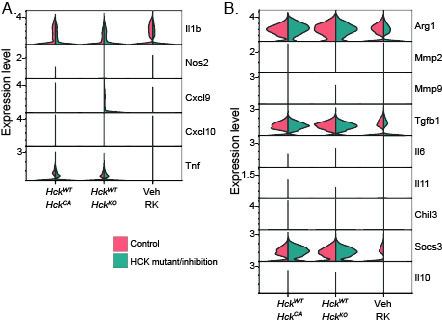
To examine the role of HCK in the differentiation of TAM phenotypes, we reclustered CD45+ immune cells (Fig. 5A). We and others have devised a consensus classification of macrophage subtypes based on single cell data across a range of tumor types to distinguish between angiogenic, immunoregulatory, interferon (IFN)-primed, inflammatory, lipid-associated, tissue resident and proliferating TAMs (3, 4). TAMs in Py8119 tumors consisted of a large immunoregulatory cluster (> 40%, folate receptor beta (Folr2)high, stabilin (Stab)1high, F13a1high), an inflammatory cluster (~ 20%, H2-Aahigh, NLRP3high, Il1bhigh) and smaller clusters of IFN-primed (~ 5%, Isg15 ubiquitin like modifier (Isg15)high, Cd274high, Cxcl9), angiogenic (~ 4%, heme oxygenase (Hmox) 1high, Vegfahigh), and tissue resident (~ 1%, lymphatic vessel endothelial hyaluronan receptor (Lyve)-1high) macrophages, with proliferating TAMs (Mki67+) accounting for 20% (Fig. 5A,B). All Py8119 tumor TAMs showed prominent expression of apolipoprotein E (ApoE)high, owing to their mammary fat pad origin, and expressed triggering receptor expressed on myeloid cells (TREM2)+, secreted phosphoprotein 1 (Spp1)high), and C1q (Fig. 5C). Finally, HCK was expressed across all TAM subtypes (Fig. 5B), and loss of HCK activity did not induce an inflammatory phenotype in any TAM clusters as determined by lack of expression of inducible nitric oxide synthase (Nos2)+ and other classically activated macrophage markers (Supplementary Fig. 4A). Similarly, there was no induction of IL-10, matrix metalloproteinase (Mmp)2 or Mmp9 expression or changes in arginase (Arg)1 expression to indicate a skewing of TAMs towards an alternatively activated phenotype, as we showed previously in several gastrointestinal tumor models (Supplementary Fig. 4B) (13–15).
Gene ontology (GO) analysis of TAM DEGs revealed that metabolic pathways were the most significantly downregulated pathways in HckKO TAMs while cytokine production and response pathways were downregulated in RK20449-treated TAMs with cell migration pathways enriched in HckCA TAMs (Supplementary File 2). However, no consistent pattern of gene expression changes emerged that could be attributed to HCK activity. Overall, scRNA-seq revealed that TAMs comprised almost half the Py8119 tumor mass and differentiated into 5 main subtypes, the proportions of which were unaffected by HCK activity. Similarly, while GO analysis results were consistent with the regulation of
Supplementary Fig. 4. HCK does not induce a classically activated or an alternatively activated TAM phenotype in Py8119 mammary tumors. (A) Violin plots comparing expression levels of classical activation markers in Py8119 tumor TAMs from control (salmon pink) or HCK mutant/inhibited (teal) mice. (B) Violin plots comparing expression levels of alternative activation markers in Py8119 tumor TAMs from control or HCK mutant/inhibited mice.
macrophage motility by HCK, no clear additional mechanism for the striking effects of HCK activity on tumor growth was revealed. This implicates the loss of HCK-activated phosphotyrosine-based motility signaling downstream of the CSF-1R in all TAM subtypes as the most likely cause.
{kind=link}
{kind=link}
{kind=link}
{kind=link}