From this study, the intertidal coral reef environments of Ryukyu Islands harbored a higher diversity of onchidiid species than previously thought (Takagi et al. 2018; Goulding et al. 2022). The high species diversity of Onchidiidae in this region may be due to the influence of Kuroshio Current in their larval dispersal, as well as unique characteristics of the Ryukyu Islands’ humid subtropical climate, different from other subtropical regions (Miyagi 1992). These conditions are likely suitable for Onchidiidae to inhabit intertidal coral reef environments.
Species diversity of Onchidiidae in intertidal coral reef environments of the Ryukyus
Genus Peronia
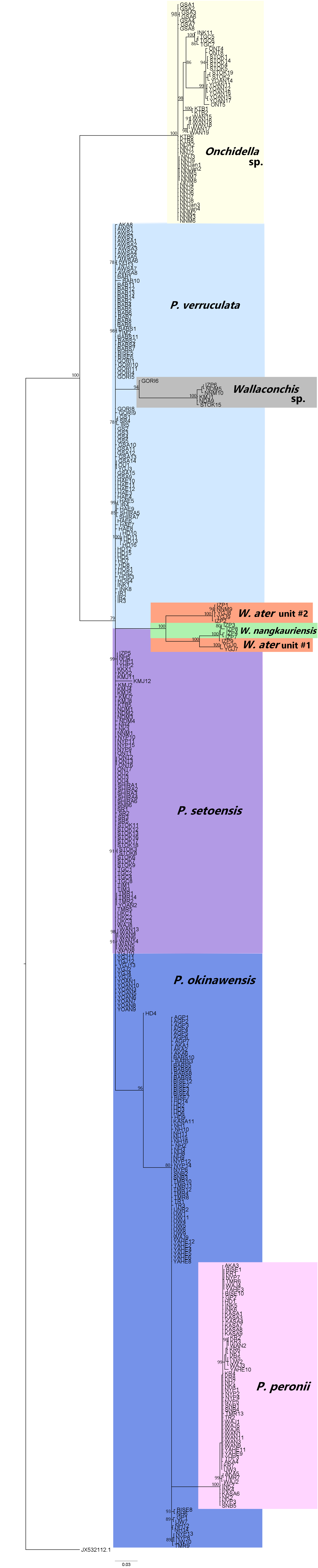
Peronia spp. are air-breathing sea slugs inhabiting intertidal reef flats, especially on hard substrates (Hamaguchi and Yoshioka 2002; Dayrat et al. 2020; Goulding et al. 2022). Peronia spp. can be found across tropical and subtropical the Indo-Pacific, from South Africa to Hawaii (Britton 1984), and some species have been frequently observed in temperate to subtropical regions of Japan (Katagiri & Katagiri 2007; Okutani 2000, Wardiatno et al. 2015). Among family Onchidiidae only genus Peronia has branchial gills on the back of the notum, which allows easy distinction from other genera (Dayrat et al. 2009). Four species of genus Peronia have been previously recorded from around the Ryukyu Islands; P. peronii, P. verruculata, P. setoensis and P. okinawensis (Mizukami et al. 2022).
Our previous study showed that the most numerous species around Okinawa-jima was P. verruculata, with P. okinawensis the most abundant around Amami-Oshima (Mizukami et al. 2022). These past observations led to the hypothesis that P. verruculata had a comparatively higher tolerance to disturbed intertidal areas, while P. okinawensis showed a preference for pristine sites. However, this idea was based on the fact that Okinawa-jima is the most densely populated island among the Ryukyus, with an area of 1208 km2 and a population of 1.3 million people (= 1,076 people/km2, Okinawa Prefectural Office, 2024; Statistics Bureau, Japan 2023). In contrast, Amami-Oshima is listed as a UNESCO Natural World Heritage site and is much more protected as a national park (Amami-Oshima 2021). Moreover, Mizukami et al. (2022) did not observe any species other than P. verruculata in Iriomote-jima, even though the island is also registered as a UNESCO Natural World Heritage Site, with pristine coastlines (Ministry of Environment 2021). Thus, this hypothesis needed to be re-evaluated with specimens from additional islands of the Ryukyus, as well as more sampling effort across Iriomote-jima, Okinawa-jima, and Amami-Oshima islands.
Consistent with Mizukami et al. (2022), P. okinawensis was the most abundant species around many islands of the Ryukyus, namely Ie-jima, Iheya-jima, Yoron-jima islands, all of which have well-preserved intertidal areas. As well, Okinawa-jima had a large number of P. verruculata, where it was the most abundant species. Moreover, P. verruculata was the only species that inhabited all intertidal coral reef environments investigated in this study. For example, P. verruculata was found at the Awase tidal flat, a site with an ongoing (since 2002) reclamation project creating an artificial island (Okinawa Prefecture 2014). Also, unlike other Peronia species, P. verruculata was found directly on artificial structures at Gushikawa Beach, Okinawa-jima. These results further suggest that P. verruculata has a higher tolerance for degraded environments, while P. okinawensis prefers more pristine intertidal areas. However, P. okinawensis was also found at Sunabe and Baba Park, Okinawa-jima, both intertidal areas that are heavily armored and considered as medium to high impacted sites (Masucci and Reimer 2019; DiBattista et al. 2020). Additionally, although P. okinawensis was reported for the first time from Iriomote-jima in this study, the most abundant species in Iriomote-jima was still P. verruculata. Since our study indicates possible relationships between species diversity and artificial coastlines, future studies should investigate coastal development and species compositions of Peronia in the Ryukyu Islands.
There were clear differences in species compositions between sampling months, particularly for P. okinawensis and P. setoensis. In this study, P. okinawensis appeared in samples from May to December, and numbers were highest during June to September and lowest in May and December. These differences in species compositions may be considered as geographical differences and investigated sites, however, sampling in Yoan, Amami-Oshima was conducted in February 2024 and the only Peronia species collected was P. setoensis. Meanwhile, during surveys in Tomori, Amami-Oshima, in November 2021, four Peronia species were collected (Mizukami et al. 2022). The surface water temperature in Amami-Oshima Island during February was approximately 20℃, and 24℃ degrees in November. Similar findings were observed in Baba Park, Okinawa-jima, where only P. setoensis was collected in December 2021, but three species (P. verruculata, P. okinawensis, P. setoensis) were found in September 2023. The average differences between mean maximum and minimum surface water temperatures in the Ryukyu Islands are approximately 7℃ to 8℃ degrees (Iryu et al. 1995). P. setoensis was described from Wakayama, located in the temperate region of mainland Japan, and previously considered as endemic to the southern Honshu area, from Boso Peninsula to Nagasaki (Katagiri and Katagiri 2007; Ueshima 2007; Dayrat et al. 2018), supporting the idea that P. setoensis tolerates colder water than the other species (Dayrat et al. 2018). The distribution of P. setoensis ranges from Okinawa-jima to Boso Peninsula, Chiba, and P. okinawensis from Iriomote to Amami-Oshima. Our results suggest that potential seasonal influences in the behavior or active time could also play a role in the habitat suitability of genus Peronia in the Ryukyu Islands, especially for P. setoensis and P. okinawensis.
Furthermore, Dayrat et al. (2020) reported that P. peronii is a nocturnal species as fewer individuals appeared during the daytime. The highest number of P. peronii specimens collected in this study was in Kurio, Yakushima (eight specimens), followed by Kasaishi Beach, Okinoerabu-jima (seven specimens). Both collections were carried out during the lowest night tides in March and December, respectively. However, all sampling in this study was conducted based on low tides, which occur during the daytime in the summer and at night in the winter in the Ryukyu Islands, and were not conducted based on circadian rhythms. Species occurrences in the intertidal areas of the Ryukyus might also be correlated with lowest tides, as well as corresponding seasonal tide differences, and thus the circadian and tidal rhythms of each species require further study.
Genus Wallaconchis
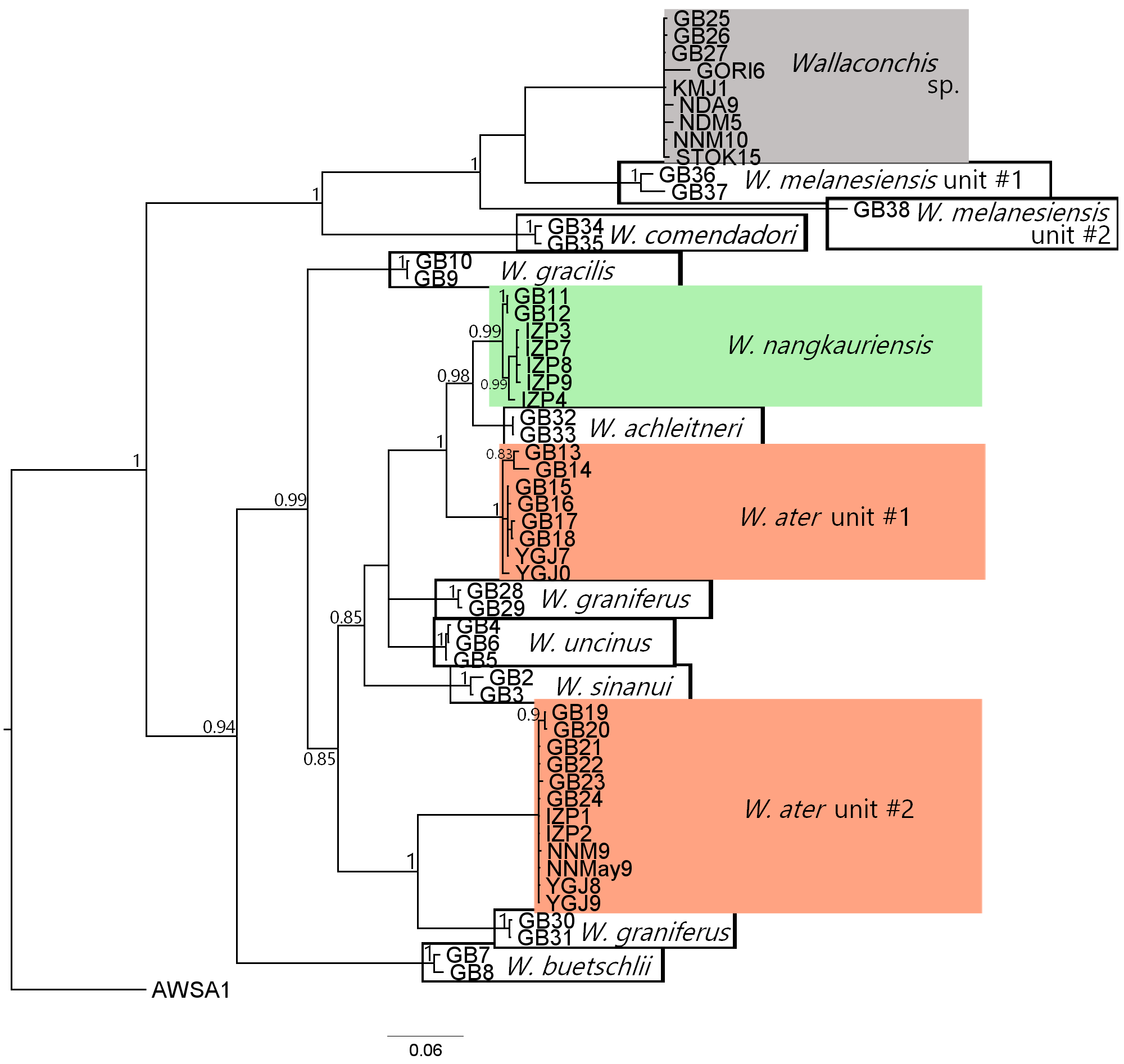
Wallaconchis is a recently described genus within Onchidiidae, and all species in this genus were originally placed in Onchidium Buchannan, 1800 (Goulding et al. 2018). Japanese literature often misidentifies this genus as genus Paraoncidium Starobogatov, 1976 with the misspelling “Paraonchidium”. According to guidebooks and literature, the Ryukyu Islands harbor three intertidal Paraoncidium species; P. fungiforme (Stantschuinsky, 1907) (Kiboshi Awamochi), P. keinense (Hoffmann, 1926) (Hime-kiboshi Awamochi), and Paraonchidium sp. (Goshiki Awamochi). The specimen pictures and descriptive anatomical information in these reports, particularly oviduct morphology, match well with the description of Wallaconchis provided by Goulding et al. (2018). Based on this information, we hereby identify Paraonchidium sp. (Goshiki awamochi) as Wallaconchis nangkauriensis and Paraonchidium keinense (Hime-kiboshi awamochi) as Wallaconchis ater (Ueshima 2014; Okutani 2017). In the description of a third species, P. fungiforme (Kiboshi Awamochi), it is mentioned that: “they inhabited at intertidal flat, the dorsal color ranges from yellow-brown to cream, with irregular black or reddish-brown spots often forming indistinct longitudinal stripes in pairs, and oviduct is narrow. Distributed in Japan to Australia” (Japanese translation of Ueshima 2014; Okutani 2017). This description does not match with any Wallaconchis (Goulding et al. 2018), and instead likely refers to Onchidina (Dayrat and Goulding 2017). However, P. fungiforme likely belongs to Wallaconchis as it co-occurred with P. keinense (= W. ater), and was mentioned as “very similar to P. keinense and the only difference was the lengths of oviduct” (Ueshima 2014).
Takagi et al. (2018) indicated that there are three species of Paraoncidium around the northern part of Okinawa-jima based on mitochondrial COI haplotype groups. Based on their registered COI sequence data in the DNA Data Bank in Japan and compared with other available GenBank sequences, these three species are likely W. ater unit #1, unit #2 and Wallaconchis sp., but no anatomical data were provided in these previous reports. Takagi et al. (2018) did not identify the different haplotype groups to species level, but found that W. ater unit #1 and #2 sympatrically existed in several intertidal areas, while Wallaconchis sp. was solely observed from only one site.
Furthermore, accurate identification of Wallaconchis species requires not only mitochondrial DNA, but also nuclear loci and anatomical information as incongruences have been observed in their mitochondrial sequences (Goulding et al. 2018). According to mitochondrial COI and 16S sequences, two specimens in our study matched with W. ater unit #1 while five specimens matched with W. ater unit #2 (Goulding et al. 2018), with 18–19% genetic distances between these two units for both markers. Goulding et al. (2018) also observed 25.7–28.9% genetic distances between W. ater units #1 and #2 based on mitochondrial markers. However, the phylogenetic tree from nuclear ITS (ITS1 and part of 5.8S and ITS2) sequences revealed that W. ater should not be separated into two units, and instead is a single species (Goulding et al. 2018). These results were also congruent with their morphological analyses (Goulding et al. 2018). This mitochondrial sequencing incongruency also occurs in other species including W. melanesiensis and W. graniferum, and it has been hypothesized to be due to maternal inheritance of ancestral sequences (Goulding et al. 2018).
Wallaconchis sp. collected in this study from Okinawa-jima, Tokunoshima, and Kikai-jima formed a distinct clade along with sequences extracted from Takagi et al. (2018), that was separated from both W. ater and W. nangkauriensis (Fig. 3). Wallaconchis sp. is located among W. melanesiensis unit #1 and unit #2 in the phylogenetic tree based on mitochondrial markers, with 13–16% and 20–22% genetic distances between species, respectively (Supp. Figure 3). However, these results are also not reliable since Goulding et al. (2018) found that W. ater units #1 and #2 did not form a sister clade in mitochondrial markers. To definitively determine if Wallaconchis sp. is a cryptic species or an already described species, incorporating anatomical information and additional nuclear marker data are needed.
Wallaconchis spp. are often observed in inner bay areas and near river mouths on smaller rocks and rubble to muddy substrates in the Ryukyus (Ueshima 2014; Takagi et al. 2021). During this study, high numbers of W. ater and W. nangkauriensis were also collected from flat intertidal areas, particularly under rubble and small rocks, rather than coral reef intertidal areas. In the Red Data book for Okinawa (2014), P. fungiforme and P. keinense were listed as threatened vulnerable (VU) species in Okinawa-jima, especially in the Awase tidal flats (Ueshima 2014). We conducted surveys during daytime low tide in August 2020 and August 2023, and at night in December 2021 in Awase, but we did not observe any Wallaconchis species in this area. Despite the claimed conservation efforts alongside the artificial island project in this area (Okinawa Prefecture 2014), there are reports across several taxa that the biodiversity of the area has already been negatively affected (Sakai et al. 2006; Kubo 2017). Thus, it is possible that Wallaconchis spp. have been extirpated from the Awase tidal flats, or that abundances have been significantly reduced in this area since 2014. Furthermore, it is likely that higher numbers of species or higher abundances of Wallaconchis inhabit mangrove forest to tidal flats in the Ryukyus. We strongly recommend that future studies examining diversity of Wallaconchis should explore not only coral reef areas but also other types of intertidal areas such as mangrove ecosystems.
Genus Onchidella
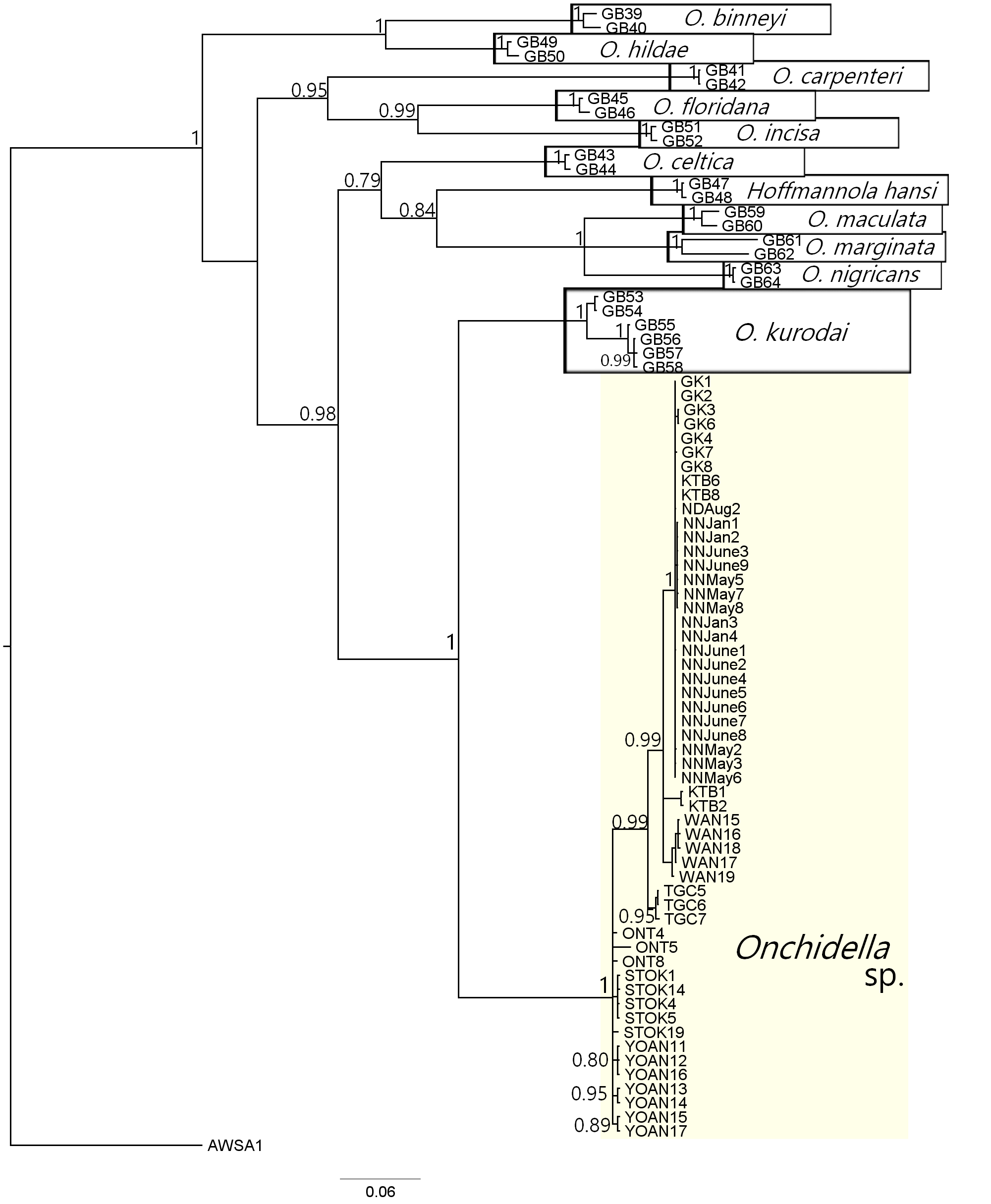
Onchidella belongs to subfamily Onchidellinae Labbe, 1934 within family Onchidiidae (Goulding et al. 2022). The distinct characteristics of Onchidella are evenly distributed marginal yellow-ish patterns and lack of dorsal eyes (Dayrat et al. 2011b). Onchidella are often found in crevices of intertidal coral reef areas (Baba 1958; Cumming 2014), but are also found on the sponge Halichondria okadai in mainland Japan (Ohgaki et al. 2011), and on bull-kelp Durvillaea spp. in the southern hemisphere (Cumming 2014).
Two species of Onchidella from Japan were described by Taki (1935); O. kurodai Iw. Taki, 1935 (Hime Awamochi) from Gogoshima and Minatoyama, Ehime, and O. orientalis Iw. Taki, 1935 (Yamato Hime Awamochi), from Tonda-mura, Wakayama. The gross morphological differences between these two species are that O. orientalis is more elongated than O. kurodai but slightly smaller, the minute papillae on O. orientalis are much numerous than O. kurodai, and that there are grey round patches on the dorsal side of O. orientalis that vary in size, which are not observed in O. kurodai.
In the Ryukyu Islands, the first observations of O. orientalis were reported from intertidal coral reef areas of Yoan, Amami-Oshima and Nakanoshima Island, Tokara (Baba 1958; Taki 1959). However, no photographic records were given, and identification criteria were not specifically mentioned for specimens. Additional reports were made for both O. kurodai and O. orientalis from mainland Japan, south of the Boso Peninsula to Ariake Bay (e.g. Nishimura 1992; Okutani and Soyama 1994; Okutani 2000, 2017; Takagi et al. 2019), O. orientalis in northern Taiwan (Cheng et al. 2005)d kurodai in Jindo Island, South Korea (Kil et al. 2005).
In Okutani and Soyama (1994) and Okutani (2000, plate 404, O. orientalis), an image of O. orientalis from Kushimoto, Wakayama includes both O. kurodai and O. orientalis, based on dorsal patterns described by Taki (1935). Later, Okutani (2017) mentioned that these two species are likely to be the same species. Recently, O. orientalis was synonymized with O. kurodai due to no anatomical differences and little genetic variation in mitochondrial COI, 16S, and 12S, and nuclear ITS2, 28S and H3 DNA sequences observed in specimens collected from both type localities (Goulding et al. 2022). However, Goulding et al. (2022) examined only three specimens of O. kurodai and O. orientalis from mainland Japan. Moreover, specimens from the Ryukyu Islands were missing from Goulding et al. (2022), and it was mentioned that Onchidella spp. are largely absent from subtropical and tropical Indo-West Pacific regions (Goulding et al. 2022). Thus, the newly collected specimens from the Ryukyus are particularly important to understand Onchidella species diversity in Japan.
Genetic distances between Onchidella specimens collected in this study from the Ryukyus and O. kurodai (Takagi et al. 2018; Goulding et al. 2022) were 19–20%, and Onchidella sp. formed a separate clade distinct from the other previously described species of Onchidella (SFig. 4). The average size of Onchidella sp. was smaller than as described for O. kurodai and is more similar to the size of O. orientalis in Taki (1935). Dorsal patterns with grey round patches were often present as in O. orientalis, but varied among individuals and some specimens did not have any patterns. Further studies with anatomical information and more genetic markers are needed to determine whether O. orientalis needs to be resurrected, or our Onchidella sp. instead represents a cryptic, undescribed species.
Onchidella spp. smaller sizes often make them challenging to notice in the field. Thus, despite our focus on this group, we feel it likely that this study overlooked Onchidella spp. at many intertidal areas, and we expect them to have much wider distribution ranges within the Ryukyu Islands than has been noted here and in previous research.
{kind=link}
{kind=link}
{kind=link}
{kind=link}