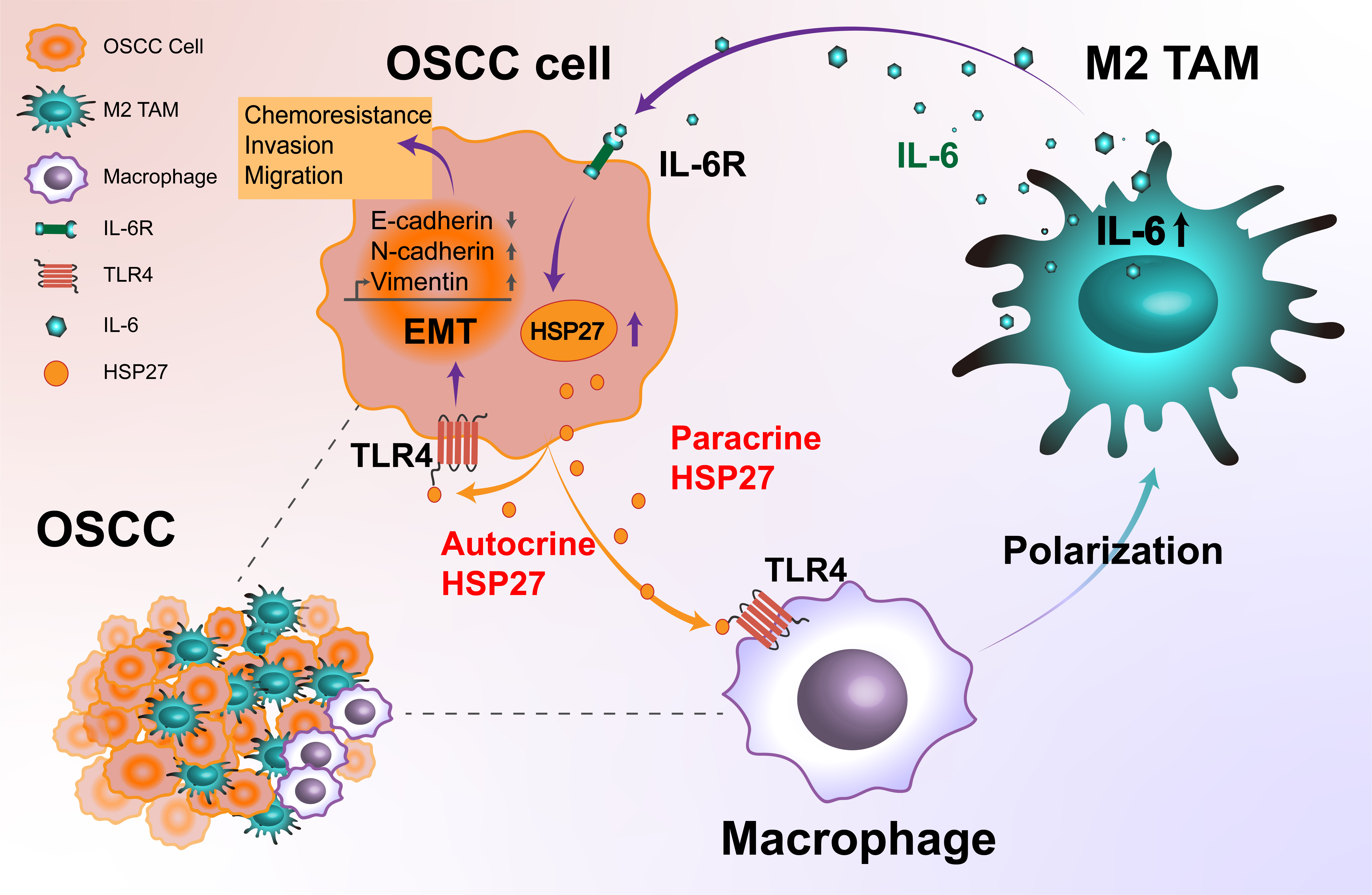
The mechanisms of interaction and crosstalk between tumor cells and TAMs have provided novel options for intervening in tumor progression [19–20]. Here, we demonstrated a higher expression of HSP27 in CAL27 cells compared with Hacat cells and TAMs. TAMs-CM significantly upregulated the HSP27 mRNA expression in CAL27 cells. Tumor-cell-derived paracrine HSP27 promoted OSCC invasion and migration through enhancing the EMT of tumor cells via binding to TLR4 on the surfaces of the TAMs. HSP27/TLR4 induced the polarization of the TAMs towards an M2-like phenotype, as well as the secretion of IL-6 in the TAMs. TAM-derived IL-6 enhanced OSCC invasion and migration via autocrine HSP27/TLR4 signaling in the tumor cells. IL-6 promoted the EMT of tumor cells via autocrine HSP27. Our study identified a novel loop of tumor cells interacting with tumor-associated macrophages in OSCC to mediate drug resistance. On the one hand, tumor-associated macrophages in the tumor microenvironment can alter both the intracellular and extracellular levels of HSP27 in tumor cells to promote their own drug resistance, on the other hand, intracellular and extracellular HSP27 from tumor cells can induce tumor-associated macrophages to secrete the cytokine IL-6, which can inversely modulate the chemoresistance to cisplatin in tumor cells, and at the same time this loop could also further enhance the stem cell properties of tumor cells, thus promoting OSCC tumor progression.
A previous study showed statistically significant differences in HSP27 expression between normal mucosa and well-differentiated OSCC cells, indicating that the expression level of HSP27 is closely associated with OSCC progression [21]. In this study, the higher expression of HSP27 in CAL27 cells compared to that of Hacat cells and TAMs further confirmed the findings of previous studies [14, 15, 33], suggesting that tumor-cell-derived HSP27 may play an important role in OSCC progression. It is noteworthy that TAMs-CM significantly upregulated the mRNA expression of HSP27 in CAL27 cells. Meanwhile, HSP27 was highly expressed in CAL27 cells, but not in TAMs. These results suggest that the highly expressed HSP27 in tumor cells may play an important role in the interaction between tumor cells and TAMs, which may be closely related to the invasion and migration of OSCC [22–23].
Previous studies have proven that tumor-cell-derived proteins may promote tumor invasion and migration by regulating TAMs [24–27]. Considering the importance of EMT in OSCC invasion and migration [28], we investigated the intervention of paracrine HSP27 from tumor cells on the invasion and migration of OSCC and the EMT of tumor cells through TAMs. Our results showed that the paracrine HSP27 from tumor cells promoted OSCC invasion and migration through enhancing the EMT of tumor cells via the TAMs. Despite there were the small changes of EMT proteins in mRNA and protein expression, they can facilitate invasion and migration to some extent. In addition, tumor invasion and migration are not only related to EMT, they may also be influenced by other factors, such as cell morphology and motility.
HSP27 also plays an important role in inducing chemotherapy resistance. In non-small cell lung cancer, specific inhibitors targeting HSP27 reduced resistance due to epidermal growth factor receptor (EGFR) to its inhibitors (TKIs)[29]. Heat shock protein αB-Crystallin with HSP27 protects cisplatin-resistant ovarian cancer cells against apoptosis induced by withaferin A and induces differentiation towards a more metastatic phenotype [30]. HSP27 was shown to be highly expressed in drug-resistant cell lines of oral squamous carcinoma, indicating that HSP27 may play a role in tumor progression and chemotherapy resistance in oral squamous carcinoma. Extracellular and intracellular HSP27 may synergistically enhance chemoresistance in tongue squamous cell carcinoma by enhancing TLR5/NF-κB signaling and interacting with BAX and BIM to inhibit the mitochondrial apoptotic pathway[31].In addition, HSP27 in the tumor microenvironment can interact with other cells in the tumor stroma to influence tumor progression. Extracellular HSP27 can upregulate the level of VEGF secreted in endothelial cells through TLR3, which in turn promotes angiogenesis [32]; extracellular HSP27 also exhibits regulatory effects on monocytes, enhances the secretion of cytokines IL-10 and TNF-α [33], blocks the differentiation of monocytes from normal dendritic cells [34] and promotes the differentiation of monocytes from tumor-associated macrophage differentiation to promote tumor progression [35]. Previous studies by our group demonstrated that OSCC tumor cell-derived HSP27 induces the conversion of TAMs to a pro-tumorigenic phenotype via TLR4 and upregulates the secretion level of IL-6, which in turn promotes the invasive migratory ability of tumor cells.
TLR4, as a main pattern recognition receptor, has been the center of scientific attention due to its contributory role in many inflammatory diseases, including septic shock and asthma. This receptor is expressed on the surfaces of a variety of cells, including macrophages, mast cells, dendritic cells, and neutrophils [36]. It can recognize a series of exogenous and endogenous danger-associated molecular patterns (DAMPs) encompassing a great variety of molecules, including HSP27. Extracellular HSP27 may activate the NF-κB signaling pathway through the TLR4 receptor on the surfaces of macrophages and promote the secretion of GM-CSF, IL-10, and IL-4 [37]. Recently, abundant evidence has shown that TLR4 on the surfaces of TAMs is closely associated with tumorigenesis and tumor progression [38–41]. There is a possibility that tumor-cell-derived HSP27 promotes the invasion and migration of OSCC and the EMT of tumor cells by binding to TLR4 on the surfaces of TAMs. Through immunofluorescence staining, our study revealed that HSP27 and TLR4 were collocated on the surfaces of TAMs. The TLR4 inhibitor TAK‐242 suppressed OSCC invasion and migration through the inhibition of EMT.
tumor-associated macrophages with a pro-tumor phenotype not only influence the proliferation and growth of tumor cells and promote angiogenesis and tissue repair, but also often secrete cytokines and specific proteins to effectively improve tumor tolerance in the presence of chemotherapeutic agents. It has been demonstrated that monocyte-derived IL-1β can generate chemotherapy resistance in pancreatic cancer cells by upregulating cyclooxygenase-2 [42]. The exosome miR-223 derived from M2-type macrophages not only reduced the apoptosis rate of ovarian cancer cells SKOV3 induced by cisplatin/paclitaxel combination, but also showed a significant positive correlation with the recurrence rate and poor prognosis of ovarian cancer patients, and has now become a recurrence and prognostic marker with outstanding potential in this field [43]. Studies have shown that multidrug resistance protein 1 (MDR1) is usually up-regulated in drug-resistant tumor cells, and macrophages in the colorectal cancer microenvironment can induce MDR1 expression through the secretion of IL-6 [44], demonstrating that TAMs-derived IL-6 possesses a complex and multidimensional regulatory mechanism for multidrug resistance (MDR).IL-6 not only regulates multidrug resistance (MDR) through the activation of several pathways, including MDR1 and BCL − 2 through the activation of several pathways and genes including MDR1 and BCL1 to promote the upregulation of their autocrine levels, thereby rendering ovarian cancer cells chemoresistant, but also by inducing the expression of C/EBP and MDR1 to produce MDR effects in breast cancer cells [45].Notably, our results indicated that OSCC tumor cells orchestrates IL-6 expression in tumor conditioned macrophages. The link between tumor cells and TAMs in OSCC chemoresistance has not been revealed before.
The interaction and crosstalk between the tumor cells and TAMs for invasion and migration in OSCC show similar regulatory mechanisms as those in other tumor types. Tumor-cell-derived mediators including RNA and proteins can promote OSCC invasion and migration through inducing the M2 polarization of TAMs. Tumor-cell-derived ANLN-210 mRNA can be transferred to macrophages via exosomes by binding to the RNA-binding protein hnRNPC and induce M2-like macrophage polarization via the PTEN/PI3K/Akt signaling pathway, thus promoting tumor proliferation, migration, and invasion of head and neck squamous cell carcinoma in a positive feedback loop [46]. A new study has shown that the PAI-1 and IL-8 produced by OSCC cancer cells contribute to the differentiation of monocytes to CD206 + TAMs [47]. In the present study, HSP27/TLR4 induced the polarization of TAMs to an M2-like phenotype, suggesting that the HSP27/TLR4-mediated OSCC invasion and migration was due to the tumor-promoting properties of M2-type TAMs.
Tumor-cell-derived mediators may also promote tumor invasion and metastasis through inducing the secretion of IL-6 in TAMs [46, 47]. The expression of hypoxia-induced high-mobility group box 1 (HMGB1) in tumor cells promoted hepatocellular carcinoma (HCC) invasiveness and metastasis via enhancing macrophage-derived IL-6 [10]. The tumor-cell-derived phosphatase of regenerating liver-3 (PRL-3) and small extracellular vesicles (SEVs) upregulated the expression of IL-6 in TAMs by activating the MAPK and TLR4/NF-κB pathways [8, 43]. In turn, IL-6 secreted from the TAMs induced the EMT program to enhance colorectal cancer invasion and metastasis via the IL-6R/STAT3 signaling pathway [8, 48]. However, the role of TAM-derived IL-6 in the interaction between TAMs and OSCC cells, as well as in OSCC invasion and migration and its underlying mechanisms, have not been completely elucidated. It was reported that the M2-type TAM-derived IL-6 promoted the PLOD2-integrinβ1axis-enhanced invasion and metastasis of OSCC [49]. The latest research has shown that the tumor-cell-derived paracrine THBS1 promotes the EMT, invasion, and migration of OSCC cells through inducing the abundant secretion of IL-6 from M1-like TAMs and the subsequent activation of the IL-6/STAT3 signaling pathway in OSCC cells. Our results proved that tumor-cell-derived paracrine HSP27/TLR4 signaling promoted the EMT, invasion, and migration of OSCC cells through enhancing the secretion of IL-6 from M2-type TAMs and the subsequent activation of the IL-6/IL-6R signaling pathway in OSCC cells. It is noteworthy that the abundant secretion of IL-6 in this study derived from M2-type TAMs, rather than M1-type TAMs, which is consistent with previous studies [50, 51, 52].
In the present study, TAM-derived IL-6 was proven to enhance OSCC invasion and migration via IL-6/IL-6R signaling. However, its underlying mechanism is not fully understood. In our previous study, it was found that the autocrine HSP27 derived from OSCC cells could promote its own invasion and migration. It was reported that tumor-initiating-cell-derived IL-33 upregulates its own expression in a positive feedback manner by increasing the TGF-βsecretion of FcεRIα + macrophages, thereby promoting the autocrine IL-33-mediated invasion of squamous cell carcinoma [53]. We hypothesized that TAM-derived IL-6, similarly to TGF-β, may be involved in regulating the expression of HSP27 in OSCC cells. The results showed that IL-6 upregulated HSP27 expression in the CAL27 cells, revealing a potential mechanism by which the TAMs-CM upregulated the HSP27 expression in CAL27 cells. Importantly, IL-6/IL-6R promoted OSCC invasion and migration via autocrine HSP27/TLR4 signaling in tumor cells. Furthermore, IL-6/IL-6R promoted the autocrine HSP27-mediated EMT of OSCC cells. In a word, TAM-derived IL-6 promotes OSCC invasion and migration by increasing the secretion of autocrine HSP27 in tumor cells through a positive feedback loop induced by TAM-derived IL-6 and tumor-cell-derived paracrine HSP27. TAM-derived IL-6 and tumor-cell-derived paracrine HSP27 forms a positive feedback loop that continuously increases the secretion of autocrine HSP27 in tumor cells. Subsequently, tumor-cell-derived autocrine HSP27 promotes OSCC invasion and migration.
CD44 and OCT4, which are common surface markers of tumor stem cell properties, also show a trend of high expression in OSCC tumor cells [54] and are now recognized to play an important role in tumor chemoresistance. It has been shown that head and neck squamous carcinoma cells, as a cell population with stem cell properties, show a trend of high expression of surface proteins (e.g., CD10, CD44, CD133), pluripotency-associated transcription factors (SOX2, OCT4, NANOG), and aldehyde dehydrogenase (ALDH) [55], whereas chemotherapy per se, i.e., chemotherapy, increases the proportion of cells with stem cell properties. Importantly, the intra-tumor heterogeneity of stem cell subpopulations also reflects the plasticity of the tumor cells, which is important for generating chemoresistance. The results of the present experiments complemented the demonstration that TAMs-derived IL-6 significantly up-regulated the expression of stem cell characteristic markers CD44 and OCT4 in two OSCC tumor cells while promoting chemoresistance, suggesting that TAMs have a positive inducing effect on the stem cell characteristics of tumors. The stem cell properties of tumors are considered to be the origin of tumorigenesis, metastasis, drug resistance and recurrence, and these two stem cell property markers may play a potentially important role in regulating chemoresistance in tumors, and we hypothesized that IL-6 may mediate chemoresistance by recognizing the potential ligand of CD44 and activating the corresponding kinases and transcription factors of stemness. Previous studies have demonstrated that the interaction between GPNMB/CD44 proteins can promote the EMT process in OSCC [56], so whether HSP27/IL-6 can also mediate chemoresistance by regulating the development of tumor stem cell properties through cell-cell interactions, and whether we can find other ligands for CD44 in this process, will also broaden the idea and direction of the design and conduct of our subsequent studies. This will also broaden our ideas and directions for the design and conduct of subsequent studies.
In summary, the present study elucidated that in the OSCC tumor microenvironment, both exogenous and endogenous HSP27 from OSCC cells can upregulate the pro-resistance effect of TAMs and promote the expression of the cytokine IL-6 by acting on TAMs, thus mediating its own chemoresistance, and through further studies, it was found that IL-6 derived from TAMs can also be reversed by HSP27 expressed by OSCC cells to promote its chemoresistance. This positive feedback loop elucidates the mechanism by which HSP27/IL-6 promotes chemoresistance in OSCC through tumor-associated macrophages, and provides a possible therapeutic target for the treatment of OSCC.
{kind=link}