In 2022, over 249 million cases of malaria were reported, with the majority of cases located in Africa (1). Efforts to control this disease, which impacts half of the world’s population, have reached a critical point in the last few years following the ten percent increase in cases observed in 2020 (2). Most of these cases are due to the unicellular eukaryotic parasite Plasmodium falciparum adapted to spread through Anopheles gambiae sl. mosquitos present throughout most of the African continent (3). Most recently, Anopheles stephensi, common to South Asia and the Middle East, has invaded the Horn of Africa and several other African countries further exasperating malaria control (4–6). The vectorial competence, breeding habitats and behaviors vary even among the members of the native An. gambiae sensu lato complex (7). With the invasion of An. stephensi now contributing to increased complexity of the already diverse vector species composition in Africa, it is important to determine the new transmission dynamics in An. stephensi invaded areas.
Interactions between Plasmodium and the mosquito midgut serve as the critical gateway for malaria transmission. The parasite invasion of the mosquito midgut requires an interaction between both parasite and mosquito proteins (8). Previous studies have shown that the level of compatibility of interacting proteins between malaria vector species and parasite species varies depending on the haplotypes of the genes coding for these proteins, especially during the midgut invasion (9). Many of the Anopheles and Plasmodium genes responsible for these interactions are currently being studied for their potential use in development of transmission blocking vaccines (10, 11).
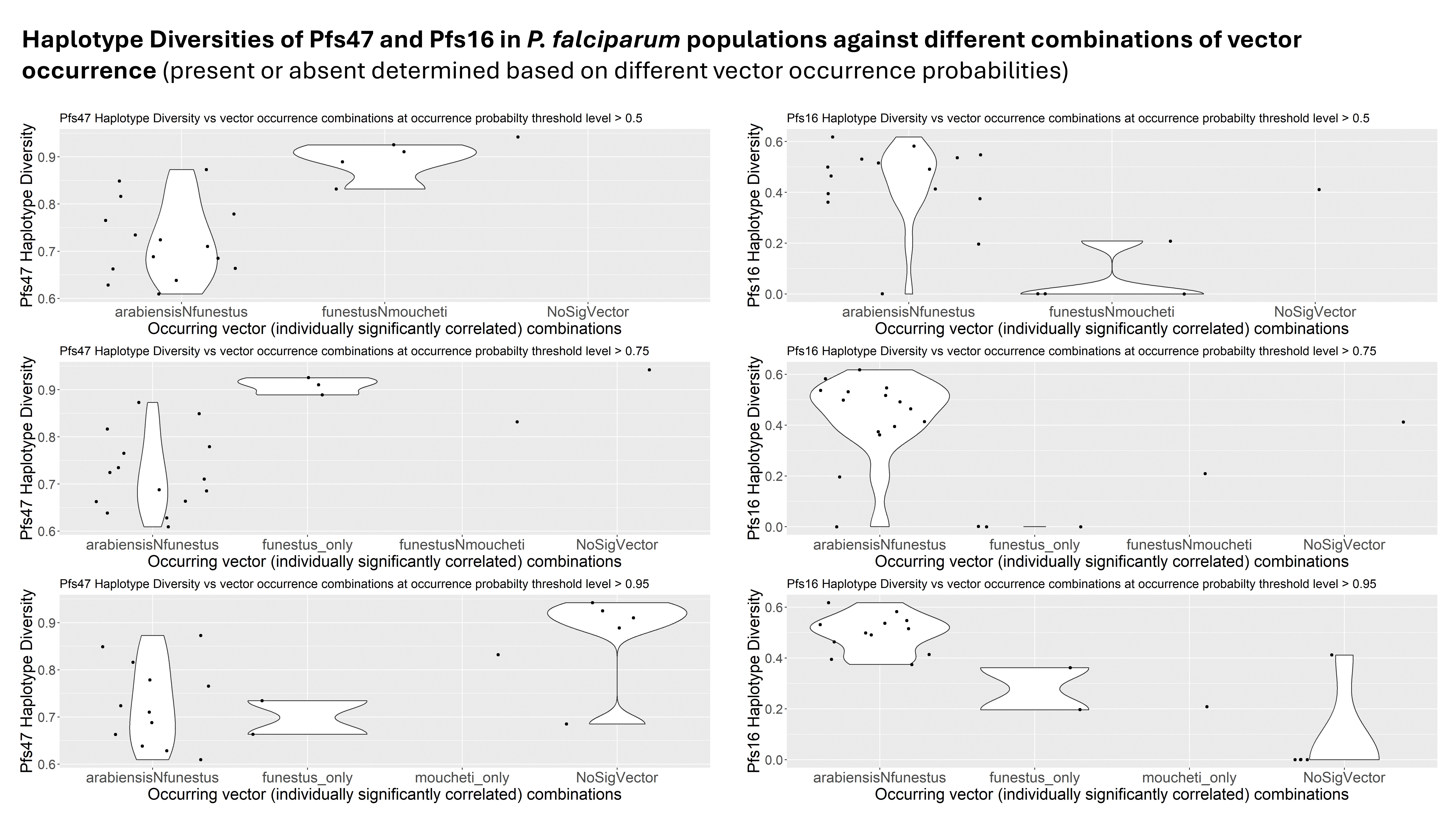
In addition to within-species genetic diversity, the haplotype diversity of genes involved in interactions with malaria parasites across different vector species may influence the genetic diversity of key genes that mediate vector-parasite interactions in Plasmodium falciparum. Different mosquito species, such as Anopheles gambiae and Anopheles funestus, have varying ecological niches, behaviors, and interactions with the malaria parasite, which can lead to differential selective pressures on the parasite's genes (12). For instance, variations in the vector's immune response, feeding habits, and geographical distribution can drive genetic diversity in the parasite as it adapts to survive and thrive in different vector environments (13). Consequently, regions with diverse vector species compositions are likely to exhibit higher nucleotide and haplotype diversity in P. falciparum genes associated with vector interactions, reflecting the parasite's adaptation to a range of vector-related selective pressures.
A well-studied example for parasite-vector interaction genes is the Pfs47-P47Rec complex in P. falciparum and An. gambiae (11, 14, 15). The mosquito midgut protein P47Rec and parasite protein Pfs47 work as a receptor-ligand pair during the Plasmodium invasion by playing a role in the immune evasion of parasites to make the parasite “undetectable” to the mosquito immune system. Silencing P47Rec expression has reduced the infection of P. falciparum in An. gambiae mosquitos (16).
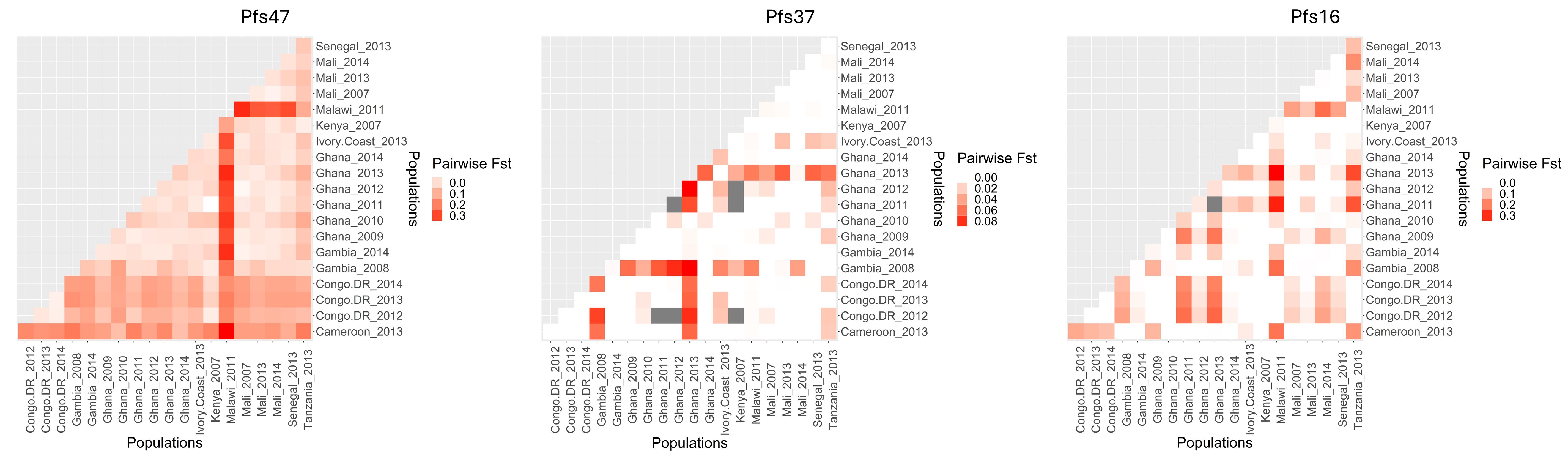
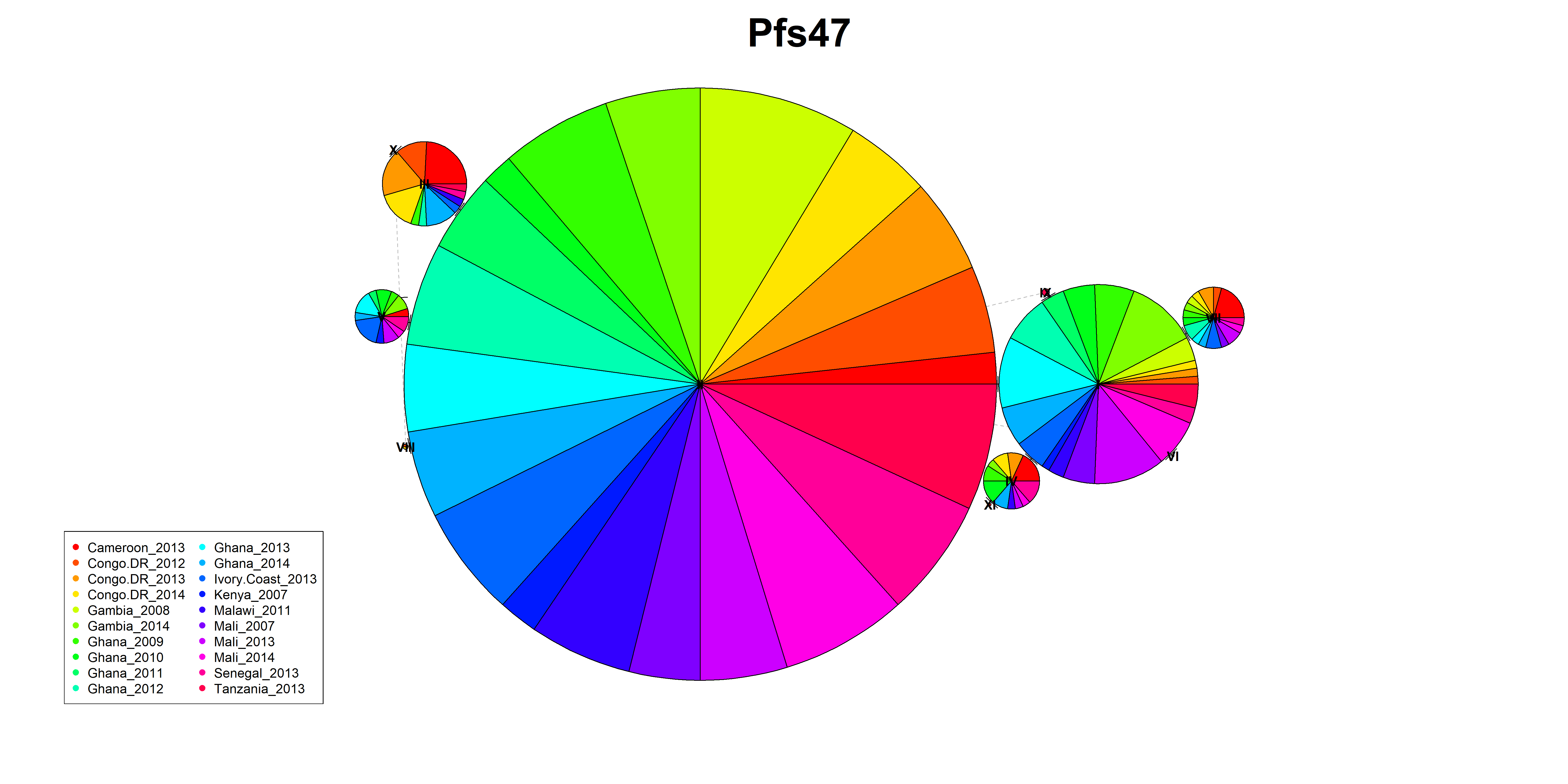
With this “lock-and-key” type mechanism, the ability of P. falciparum strains to invade the Anopheles midgut cells is dependent on the correct matching of Pfs47 surface protein haplotype (“the key”) with the Anopheles midgut receptor P47Rec (“the lock”) (16). Previous functional studies demonstrated that replacing Pfs47 haplotype in African P. falciparum with a different haplotype from another continent is sufficient to change the compatibility between the vector and parasite (9). Later studies have shown that Pfs47 is important for the adaptation of Plasmodium falciparum to different malaria vectors (17). This vector-mediated selective pressure at the continental level in Pfs47 resulted in significant population structure between different continents, particularly in domain 2 of the protein (14, 18, 19). Sub-continental selective pressure on the Pfs47 has been observed in previous studies in Nigeria, Brazil, and Malaysia (19). Still, significant knowledge gaps remain about the level of vector-mediated selective pressure on Pfs47 at a sub-continental level in Africa. This is important to evaluate given the multiple Anopheles vector species that exist sympatrically across Africa, some quite divergent from one another (eg. the An. gambiae complex vs. An. funestus) (20).
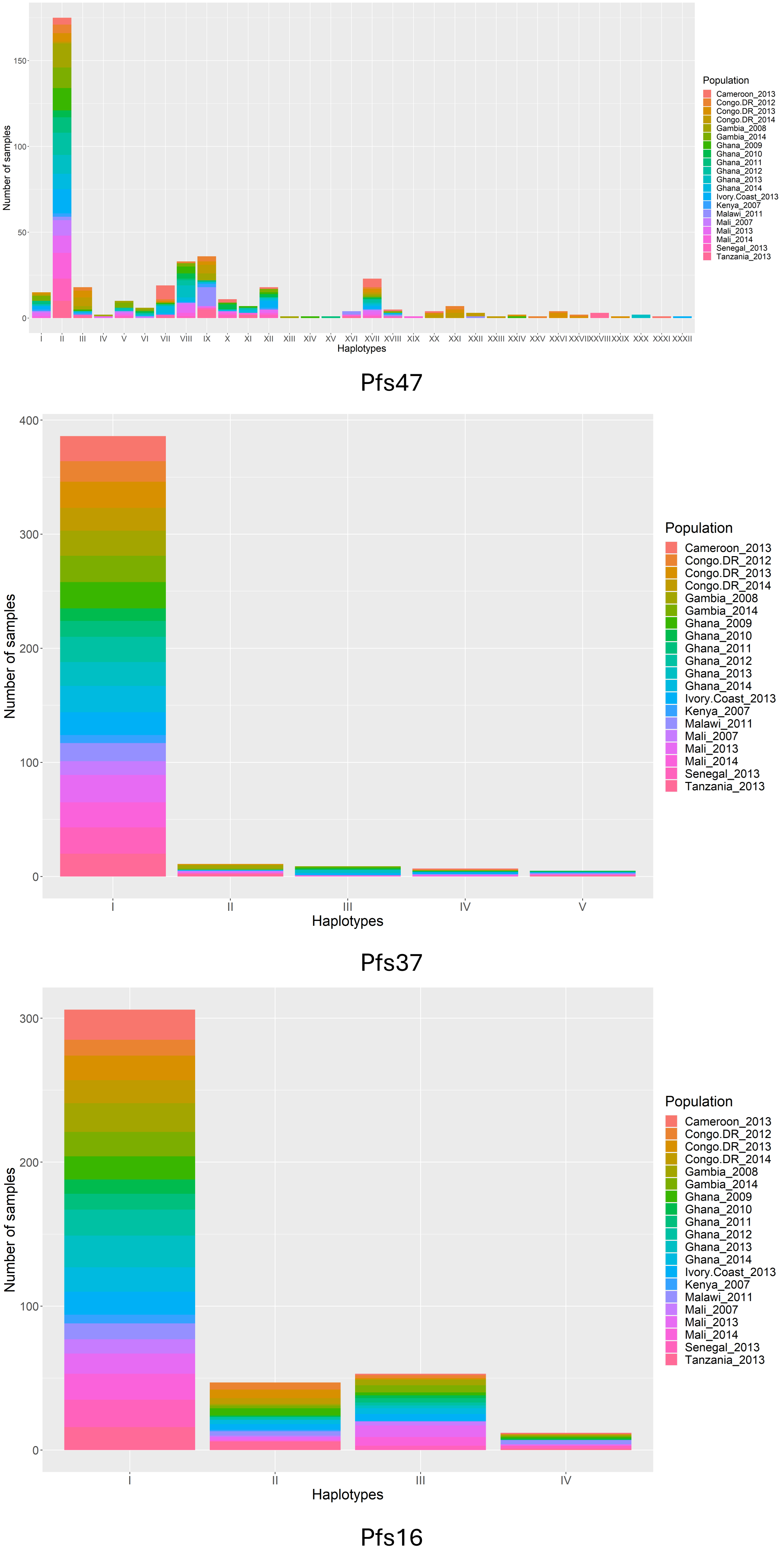
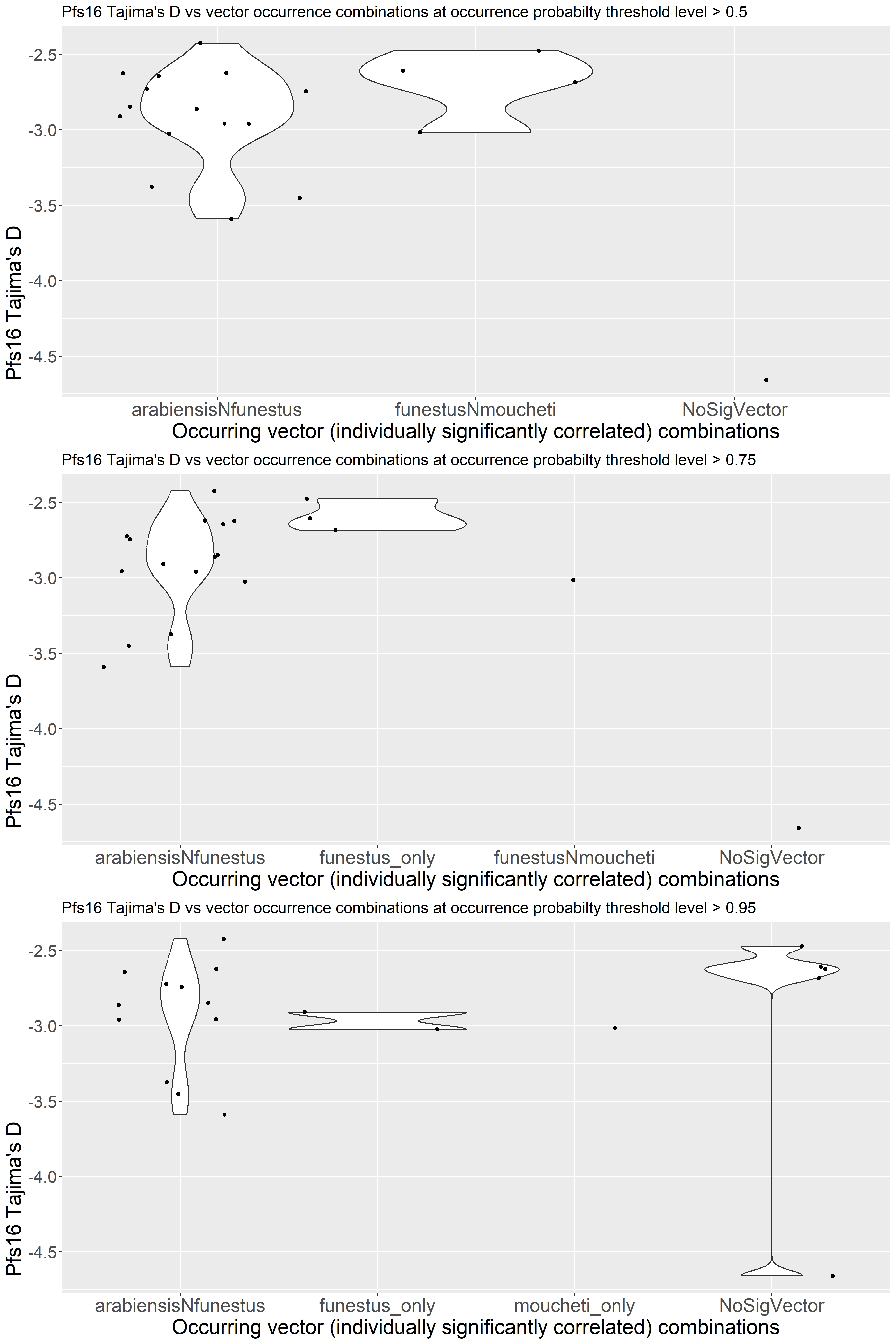
In addition to the Pfs47-P47Rec system, there are several other protein coding genes being studied as transmission blocking vaccine (TBV) targets based on their role in parasite-vector compatibility. Therefore, like Pfs47 and P47Rec, these parasite and vector genes may also involve vector-mediated selection (10, 15). In our study we selected two more genes from P. falciparum and another two genes from An. gambiae which are important for vector-parasite interactions. In Plasmodium parasites, Pfs16 (PF3D7_0406200) and Pfs37 (PF3D7_1204400) have been recognized to be important for vector-parasite interactions because of their significant upregulated expression in the sexual stages and interactions with mosquito midgut proteins (15). Knocking out Pfs16 or Pfs37 has shown a reduction in the number of oocysts generated in the mosquito midgut during the parasite invasion (15, 21). Initially Pfs16 was suspected to be required for optimal production of sexual stage parasites (22).
At the other end of the vector-parasite interaction equation, Anopheles midgut proteins AnAPN1 and HPX15 have been recognized for their importance in vector-parasite interactions and for their significant impact on the survival of the parasite. HPX15 is an immune-related protein with pattern-recognition molecules and previous studies indicate that it promotes malaria transmission (10). Specifically, HPX15 plays a role in the preservation of the functionality of stored sperm and long-term fertility in An. gambiae (23). In An. stephensi mosquitos, RNA interference-mediated silencing of midgut AsHPX15 gene has drastically reduced the number of developing P. berghei oocysts (24). Alanyl aminopeptidase N (AnAPN1) is a protein that can elicit transmission-blocking antibodies, which is believed to be highly conserved among Anopheles vectors (25), though not thoroughly investigated across Africa. The effectiveness of antibodies targeting AnAPN1 against P. falciparum and P. vivax across distantly related Anopheles species is well studied (26).
While there is strong support of ancient vector-mediated selection on Plasmodium by continentally structured Anopheles species, questions remain about the potential for ongoing vector-mediated selection within a continental region. The goal of this study was to evaluate the potential for vector-mediated selection on parasite populations within African countries by examining the patterns of diversity in vector-parasite interacting genes. To better understand the sub-continental dynamics of vector-mediated selection, we aim to investigate the genetic diversity and selection signals in P. falciparum interaction genes, along with the vector species composition and subspecies variation in An. gambiae interaction genes. This study will specifically address the implications of Anopheles stephensi invasion by providing insights into the nature of these interactions on a finer scale.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}