The tested fungi enhance biomass production in tomato under drought conditions
The effects of fungal endophytes on plants under abiotic stress are extensively studied. S. indica confers tolerance to drought in maize (Zhang et al. 2018), tomato (Azizi et al. 2021) and barley (Ghabooli et al. 2013). P. minioluteum is reported to confer salt stress tolerance in soybeans (Khan et al. 2011), and P. chrysogenum confers drought and salt stress tolerance in tomato (Morsy et al. 2020).
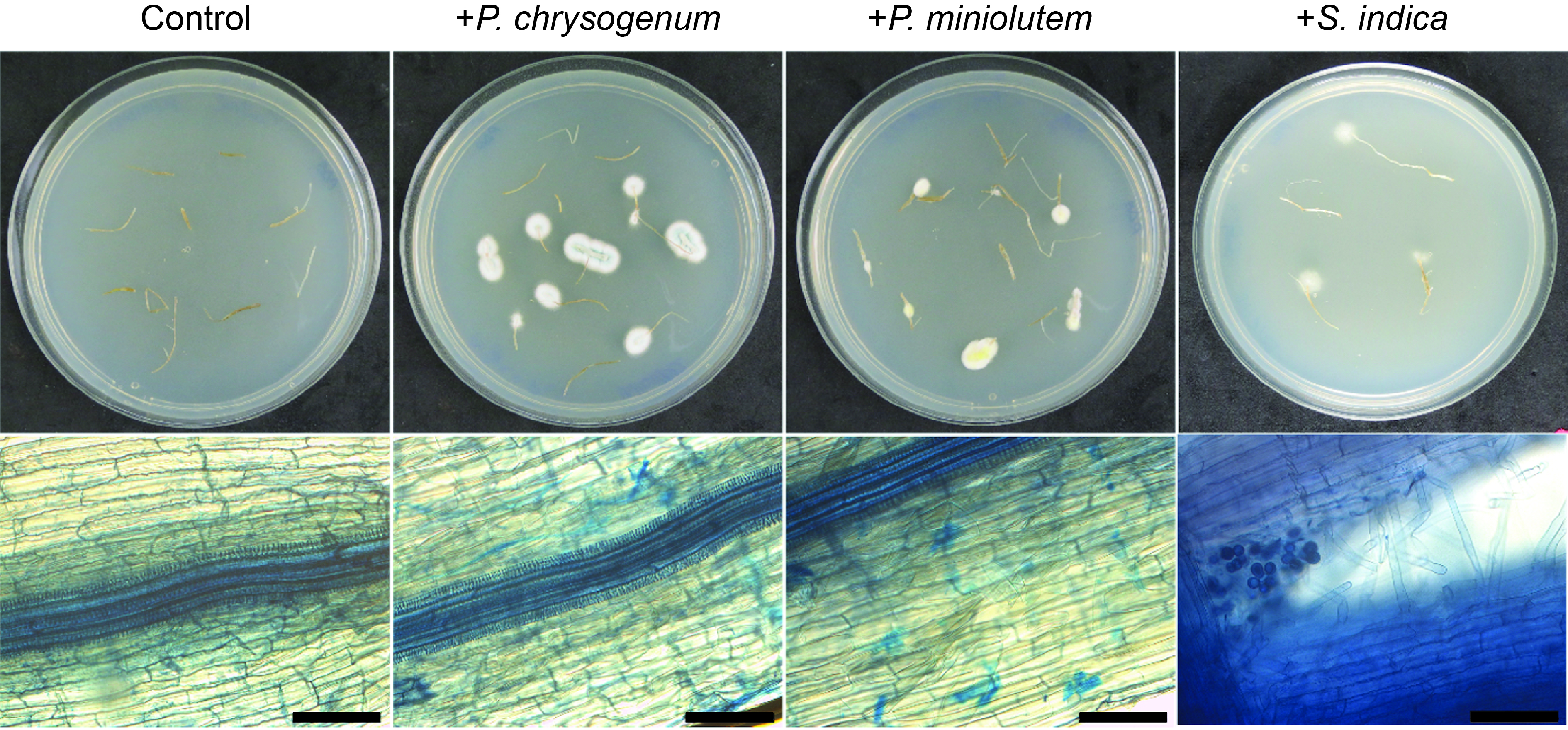
In this study, the drought stress alleviating effects of these three different fungal endophytes on S. lycopersicum plants were investigated. Initially, the capacity of the fungal endophytes to infect the tomato seedlings was assessed to ensure a comparable test system. Through re-isolation experiments and trypan blue staining, it was confirmed that S. indica, P. chrysogenum, and P. minioluteum were capable of infecting the roots of S. lycopersicum seedlings (Supplementary Fig. S1).
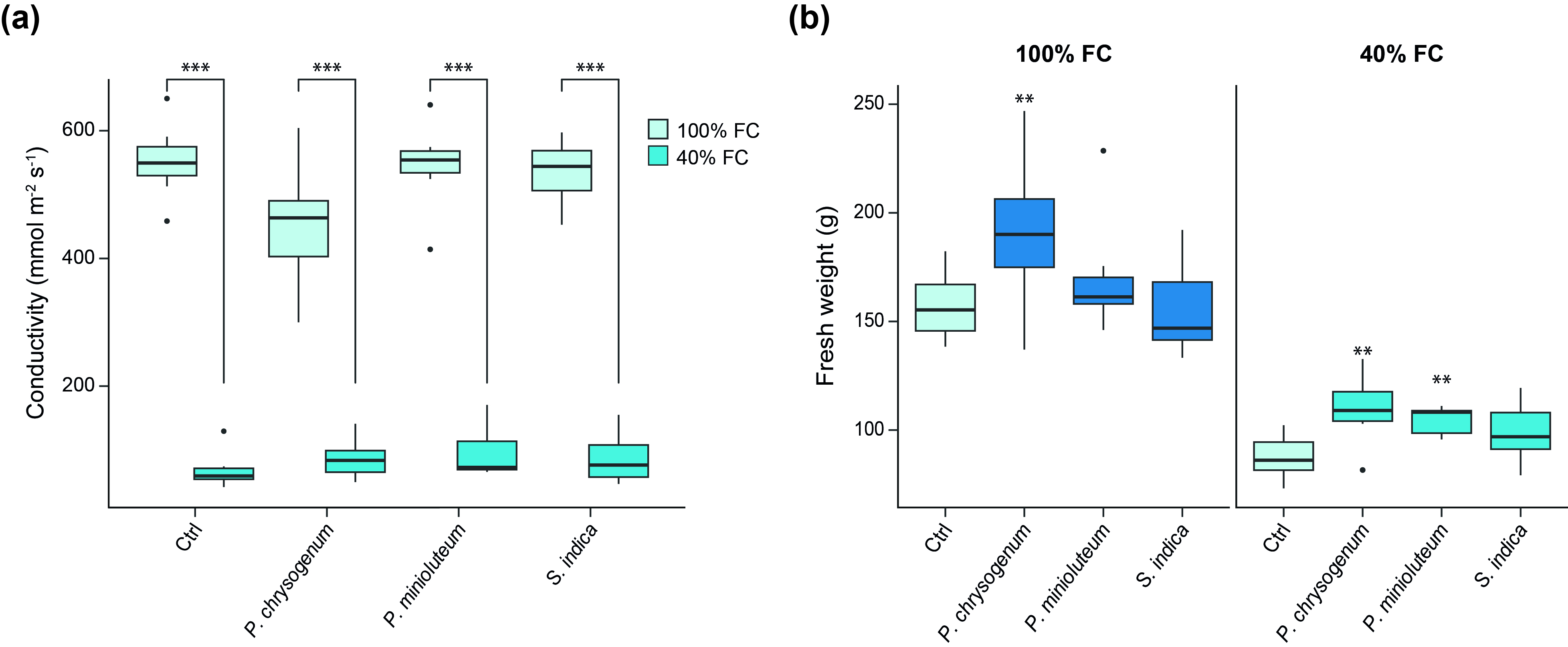
Six weeks after the second infection and drought treatment, several physiological measurements were conducted. As illustrated in Fig. 1a, significant differences in plant height were observed in watered plants inoculated with the different fungal endophytes. Notably, the values obtained under drought conditions did not exhibit significant differences. Regarding biomass, after measuring the dry weight of the aerial parts, significant differences were observed between watered plants inoculated with P. chrysogenum and plants under drought conditions inoculated with P. chrysogenum, P. minioluteum, and S. indica (Fig. 1b).
In addition to height and dry weight, stomatal conductance and fresh weight were also analyzed (Supplementary Fig. S2). Stomatal conductance was measured to verify the effects of the applied drought conditions. The applied stress conditions were confirmed, as the stomatal conductance under drought stress (40% FC) was significantly lower than under the control conditions (100% FC). Given the significant dry weight enhancement observed in the aerial parts of inoculated plants under drought conditions compared to control plants, we subsequently conducted an RNA-seq analysis to elucidate the molecular mechanism(s) underlying the observed increased biomass in endophyte-inoculated plants. Considering the root endophytic lifestyle of the three fungi, root tissue from mock-treated and fungus-infected plants was harvested and analyzed 6 weeks post-infection.
Transcriptional response to drought stress and fungal infection
Comparison of the transcriptional response of non-inoculated control plants (Ctrl40 vs Crtl100) to drought stress with the joint responses of inoculated plants (F40 vs F100) subjected to identical conditions revealed that the number of DEGs in inoculated plants is substantially reduced compared to non-inoculated plants, suggesting a potential stress-mitigating effect of the fungi. As demonstrated in Fig. 2a, a total of 4250 DEGs were identified for the control plants, whereas only 1099 DEGs were observed in plants co-cultivated with the fungi. Notably, upon applying a threshold of log2 (fold change) ≥ |1|, the number of DEGs that exhibited a response to the fungal infection under control conditions (F100 vs Ctrl100) and drought stress conditions (F40 vs Ctrl40) was relatively low, with 150 and 401 upregulated and 523 and 291 downregulated genes, respectively. The comprehensive analysis of the effects of individual inocula on tomato plants under drought conditions compared to watered reference plants (Ctrl) demonstrated that plants inoculated with P. chrysogenum exhibited the highest number of DEGs (5751), followed by plants inoculated with S. indica (5017), and P. minioluteum (4158). A principal component analysis (PCA) conducted on the normalized read counts (FPKM) of the whole RNA-seq dataset clearly illustrated the major impact of drought stress. The datasets can be categorized into two groups along the first principal component that explains 37% of the variance, distinctly separating the watered from the drought stress samples (Fig. 2b).
However, no significant difference was observed between inoculated (Pch, Pmin, and Sind) and non-inoculated samples (Ctrl). This observed lack of distinct separation among the groups may be attributed to the experimental design. In an effort to simulate agricultural field conditions, the tomato plants were cultivated in non-sterilized soil within a greenhouse environment with controlled temperature parameters. Nevertheless, this inherent variability, in conjunction with the six-week interval between root inoculation and tissue collection, may potentially contribute to the observed sample variability. However, at least under drought stress conditions, the P. chrysogenum (Pch) and S. indica (Sind) samples appear to cluster together and separate from the control plant group.
As illustrated in Fig. 2c, subsequent comparative analysis of the distinct DEG groups under drought conditions revealed the presence of a substantial cohort of DEGs (2650) shared across all inoculated conditions and the non-inoculated condition, comprising 1211 upregulated and 1439 downregulated genes, respectively. This substantial number of shared DEGs indicates a common response to drought conditions in both non-inoculated and inoculated plants. This observation suggests that co-cultivation with the investigated fungi may enhance the molecular mechanism(s) typically activated in response to drought stress. In addition, the analysis highlighted certain DEG groups exclusively shared by fungus-infected plants, which supports the hypothesis of specific responses to fungal infection. Regarding the DEGs shared under all test conditions, it is important to consider that the expression levels of these DEGs may vary significantly between inoculated and non-inoculated conditions. To address this possibility, we conducted an analysis of potential gene expression correlations under drought conditions with the aim of elucidating the mechanisms by which fungal endophytes may enhance drought tolerance in tomato plants.
Co-expression modules in tomato roots associated with drought and fungus infection
A weighted gene co-expression network analysis (WGCNA) was conducted to examine gene expression patterns under varying conditions, including the presence or absence of endophytes, and under watered or drought conditions.
The WGCNA module trait heatmap, depicted in Fig. 3, demonstrates the identification of 7 modules, each denoted by a distinct color. Correlations between modules and conditions with a p-value < 0.05 were deemed statistically significant. The green module exhibited the most significant positive correlation with P. chrysogenum (r = 0.44, p = 0.032) and S. indica (r = 0.54, p = 0.0065), while the turquoise module displayed a positive correlation with P. chrysogenum (r = 0.49, p = 0.015) and S. indica (r = 0.42, p = 0.043), and the yellow module with P. minioluteum (r = 0.51, p = 0.01). In contrast, negative correlations were observed in the blue module with P. chrysogenum (r = − 0.43, p = 0.036), and the grey module with P. chrysogenum (r = 0.58, p = 0.003).
From these correlations, we focused our attention on those that elucidated the primary shared response among the different conditions. We anticipated identifying a module with significant correlations for all tested endophytes. However, as illustrated in Fig. 3, no more than two correlations were observed within a given module. Consequently, we proceeded to examine the correlations in these modules, specifically the green and turquoise modules, and the correlations between inoculations with P. chrysogenum and S. indica. The turquoise module was ultimately selected due to its higher correlation coefficients and because it also demonstrated a tendency to exhibit a positive correlation with P. miniolutem, although this correlation was not statistically significant. Furthermore, it was hypothesized that this module contains gene correlations that could potentially elucidate the more pronounced promotion of shoot biomass production observed in tomato plants inoculated with P. chrysogenum compared to those co-cultivated with S. indica (Fig. 1b).
Extraction of genes associated with the turquoise module and the inoculated conditions with P. chrysogenum and S. indica (module eigengene-based connectivity (kME) cut off = 0.5 and gene significance (GS) cut off = 0.4) resulted in the identification of 1693 genes from P. chrysogenum-infected plants and 947 genes from S. indica-infected plants. Subsequently, these two gene groups were utilized to perform a Venn diagram analysis to identify genes at the intersection of the compared conditions. As depicted in Fig. 4a, 556 genes were found to be shared between the two conditions. To ensure the most comprehensive classification of the genes, we opted to search for the A. thaliana orthologs of the 556 tomato genes and conduct the GO enrichment analysis on the Arabidopsis orthologs. A total of 334 Arabidopsis ortholog genes were obtained and employed for the GO analysis.
The GO enrichment analysis revealed multiple overrepresented GO terms (Supplementary Table S2). The most significantly enriched GO:Biological Process (BP) terms were 'single-organism process' (GO:0044699), 'single-organism cellular process' (GO:0044763), 'single-organism metabolic process' (GO:0044710), 'small molecule metabolic process' (GO:0044281), 'metabolic process' (GO:0008152), 'response to stimulus' (GO:0050896), 'response to endogenous stimulus' (GO:0009719), 'organic substance metabolic process' (GO:0071704) and 'alpha-amino acid biosynthetic process' (GO:1901607). Following the elimination of redundant GO terms, we focused on a specific cluster of GO terms that includes 'response to hormone' (GO:0009725), 'response to water deprivation' (GO:0009414), 'response to acid chemical' (GO:0001101), 'response to abiotic stimulus' (GO:0009628), 'response to chemical' (GO:0042221) and 'defense response' (GO:0006952), among others (Fig. 4b).
Core drought stress response module identification in P. chrysogenum and S. indica inoculated roots
With the objective of identifying candidate genes responsible for the enhanced drought tolerance observed in tomato plants inoculated with P. chrysogenum and S. indica, respectively, 59 genes associated with the Gene Ontology (GO) terms 'response to hormone' (GO: 0009725), 'response to water deprivation' (GO: 0009414), and 'response to abiotic stimulus' (GO: 0009628) were selected to construct a functional interaction network. As illustrated in the heatmap (Fig. 5a), three distinct clusters of genes were identified based on their expression patterns. A group of 26 genes exhibited higher expression in CTRL plants compared to plants inoculated with the fungi. A second group, comprising four genes, demonstrated higher expression in S. indica inoculated plants compared to CTRL plants and plants co-cultivated with P. chrysogenum. Finally, a third group containing 29 genes was characterized by higher gene expression in the roots of the inoculated plants compared to CTRL plants. The presence of the two large groups supports the hypothesis of a shared drought response in plants inoculated with the two different endophytes.
In the first cluster, we identified genes associated with ABA perception and signaling, including PYRABACTIN RESISTANCE 1 (PYR1), PYR1-LIKE 4 (PYL4), and arabinogalactan protein SALT OVERLY SENSITIVE 5 (SOS5); genes related to auxin perception and signaling such as TRANSPORT INHIBITOR RESISTANT 1 (TIR1) and AUXIN RESPONSE FACTOR 4 (ARF4); transcription factors (TFs) involved in stress response, such as ETHYLENE RESPONSIVE FACTOR 062 (ERF062), ERF107, and SENSITIVE TO PROTON RHIZOTOXICITY 1 (STOP1); and two genes associated with jasmonic acid response, namely the phosphatase VEGETATIVE STORAGE PROTEIN 1 (VSP1) and a 6,7-dimethyl-8-ribityllumazine synthase known as CORONATINE INSENSITIVE1 SUPPRESSOR 1 (COS1). In addition to hormone-related genes, the first cluster comprises two genes associated with ion homeostasis and the salt stress response: SOS3, a calcium sensor essential for potassium (K+) nutrition and salt expulsion, and HIGH-AFFINITY K+ TRANSPORTER 1 (HKT1), a sodium (Na+) transporter.
In the second cluster, we identified a gene associated with auxin signaling, ARF17, and a gene related to proline biosynthesis, ∆1-PYRROLINE-5-CARBOXYLATE SYNTHASE 2 (P5CS2).
The third cluster comprises genes involved in ABA signaling, including ABA INSENSITIVE 1 (ABI1), ABI FIVE BINDING PROTEIN 1 (AFP1), AFP2, HOMOLOGY TO ABI2 (HAB2), and HIGHLY ABA-INDUCED 1 (HAI1). In addition to genes associated with ABA signaling, this cluster also contains genes related to ABA transport, such as ATP-BINDING CASETTE G25 (ABCG25), ABCG13 and ABCG22, as well as genes involved in ABA response, including OUTER MEMBRANE TRYPTOPHAN-RICH SENSORY PROTEIN (TSPO) and TFs such as G-BOX BINDING FACTOR 3 (GBF3), HOMEOBOX 7 (HB7), a NAC TF known as RESPONSIVE TO DESICCATION 26 (RD26), and a homeodomain zip (HD zip) class II TF HAT22. Furthermore, the third cluster encompassed the potassium transporter STELAR K+ OUTWARD RECTIFIER (SKOR) gene, the heat shock factor gene HEAT SHOCK TRANSCRIPTION FACTOR A2 (HSFA2), a gene associated with auxin response SMALL AUXIN UPREGULATED RNA 14 (SAUR14), and a gene involved in ethylene biosynthesis 1-AMINOCYCLOPROPANE-1-CARBOXYLIC ACID SYNTHASE 6 (ACS6). Subsequently, we utilized these genes to conduct a functional interaction network analysis to elucidate relationships among the identified genes.
The network analysis confirmed the involvement of several genes in processes related to hormone signaling and biosynthesis. The analysis of the network topology revealed five nodes with considerably high degrees of connectivity: ABI1 (12 edges), HAI1 (8 edges), HB7 (6 edges), HAB2 (5 edges), and SOS3 (4 edges) (Fig. 5b). Among these five nodes, three are protein phosphatase 2Cs (PP2Cs) and one is a TF related to ABA responses, emphasizing the significance of ABA in the drought response elicited by the inoculation of P. chrysogenum and S. indica. The network analysis (Fig. 5b) also highlighted a central core of genes that are potentially responsible for the drought stress response and increased tolerance phenotype exhibited by the plants inoculated with P. chrysogenum and S. indica. In addition to the ABA-related nodes, the network included three genes associated with ion homeostasis, namely SOS3, HKT1, and SKOR, as well as the three auxin-related genes TIR1, ARF4, and ARF17.
To validate the accuracy of our RNA-seq data, we conducted a RT-qPCR analysis for 4 of the principal genes identified in the network: HB7, RD26, ABI1, and HAI1, and assessed the correlation of the data. As illustrated in Fig. 5c, the data demonstrate substantial correlation for the selected genes under the given test conditions, which supports the robustness of the derived network. The differential expression levels of the genes are presented in Supplementary Table S3.
{kind=link}
{kind=link}