Oxidizing biocides (e.g. chlorine) have been mostly used to combat biofouling in cooling water systems worldwide. Among them chlorine has been the most preferred biocide of choice due to its low cost, ease of handling and known breakdown products (Rajagopal et al., 1995; Nair et al., 1997; Venkatnarayanan et al., 2017). Kalpakkam a rocky shore area, located on the South East coast of India, is a high biofouling potential site with heavy fouling by green mussels observed in the cooling water systems (Rajagopal et al., 1997). The green mussel Perna viridis has been the extensively studied organism with respect to fouling control using oxidizing biocides (Rajagopal et al., 2003a; Chavan et al., 2018), mortality and valve movement response (Rajagopal et al., 1996), filtration activity, byssogenesis (Rajagopal et al., 2006), antioxidant enzyme activity (López-Galindo et al., 2010), genotoxicity (Chavan et al., 2016), and physical health conditions (Masilamoni et al., 2002). The power station abstracts coastal seawater at the rate of 33 m3 sec− 1 with a velocity of ~ 3 m sec− 1 through a sub seabed tunnel, which has been heavily infested by green mussels (Perna viridis) with loading of 211 kg m− 2 observed in the vertical shaft of the seabed tunnel (Rajagopal et al., 1996). The station practices a low dose continuous chlorination regime of 0.2 ± 0.1 mg L− 1, which is otherwise called as “exomotive chlorination”, essentially meant to drive out mussel larval spat without settling inside the CWS. However, under practical circumstances in large cooling water systems, proper reach of the biocide near the wall is questionable to the varying geometries and sizes of the conduits, which results in establishment of biofouling communities. Continuous low dose chlorination with residuals of 0.2 ± 0.1 mg L− 1, has been found to be ineffective against settled adult green mussels (Masilamoni et al., 2002). Green mussels have been shown to detect chlorine concentrations as low as 0.15 mg L− 1 and complete valve closure occurs above 0.55 mg L− 1 (Masilamoni et al., 2002). It has been demonstrated using the Mossel Monitor® device, that valve closure in Perna viridis was initiated at residuals of 0.7 mg L− 1 and 100% valve closure occurs at concentration of 1.0 mg L− 1 (Venkatnarayanan et al., 2021). To combat mussel fouling in the process seawater heat exchangers (PSWHX) and their conduits and to comply with the discharge regulation and to improve the efficiency of the PSWHX supplementary biocidal addition using actibromide® has been envisaged. It is important to investigate the sub-lethal physiological responses of mussels which offer better indices than lethal responses in planning at a biocide dosing strategy. Quite often, high chlorine residuals, results in denaturation of cell membranes leading to lethal effects, particularly in the gills (Opresko, 1980), but at low residuals, other physiological activities of mussels are affected (Khalanski & Bordet, 1980). Exploring the impact of sub-lethal and lethal concentrations of continuous actibromide® on mortality and physiological changes in adult green mussels would help in fine tuning the biocidal regime to be adopted.
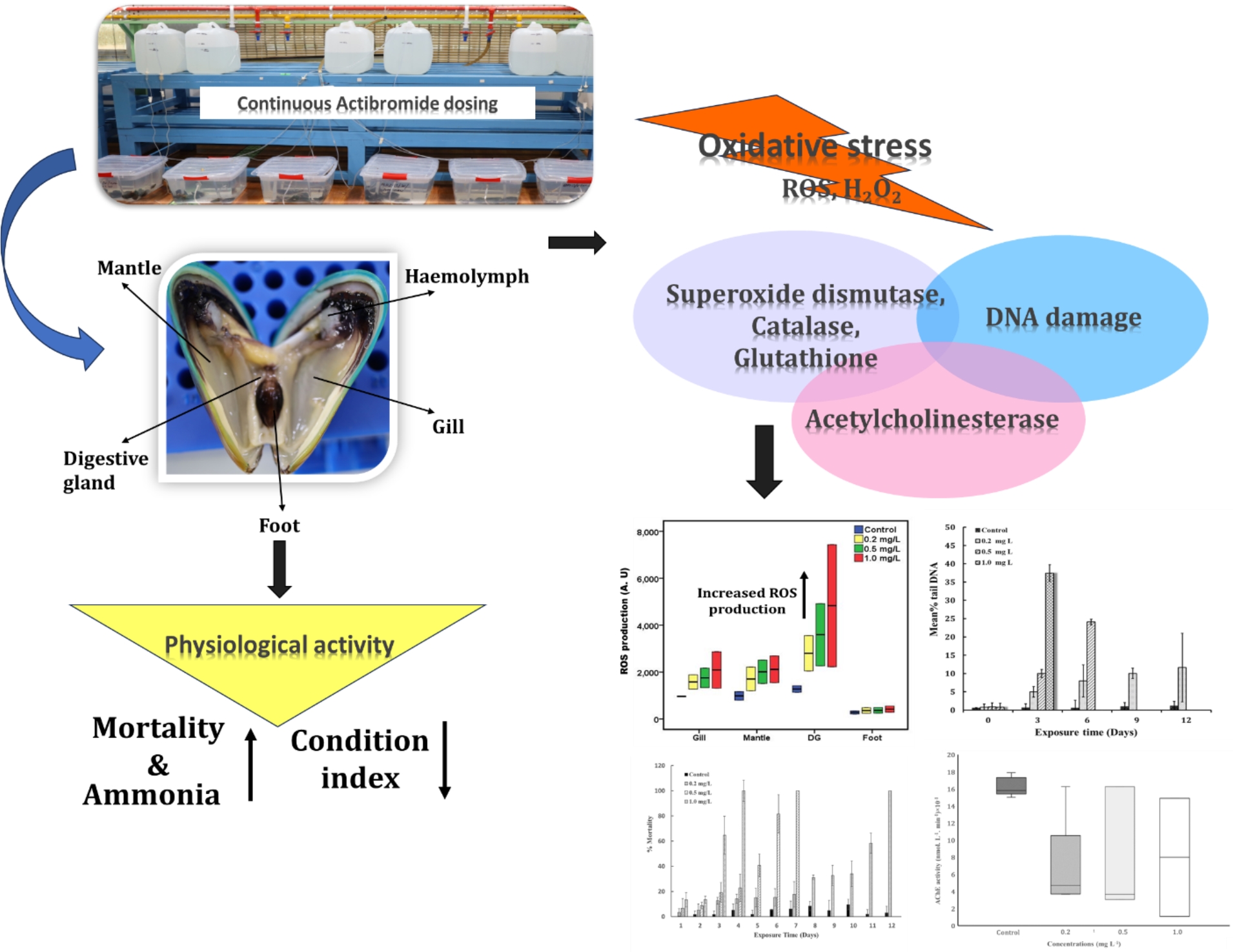
In the present study, the observed time taken for mortality for adult green mussels was found to be 12 days at 0.2 mg L− 1, 7 days at 0.5 mg L− 1 and 4 days at 1.0 mg L− 1 actibromide® residuals. Investigation showed, the 100% mortality appears to be much lower in continuous actibromide® residuals than the previously reported for chlorine for P. viridis (Rajagopal, 1995), Mytilus edulis (James, 1967), Mytilopsis leucophaeata (Rajagopal et al., 2003a), and Dreissena polymorpha (Van Benschoten et al., 1995). It has been reported that at 1.0 mg L− 1, mussels were able to open their valves to feed, although at a residue rate (White, 1966; Rajagopal et al., 1991b). Such high concentrations cannot be used practically in CWS from an environmental standpoint and biocidal discharges in marine outfall are regulated with an upper threshold limit of continuous discharge of 0.2 ± 0 actibromide®. 1 mg L− 1 (Jensen, 1982; Venugopalan et al., 2011). In the present study, an elevation in stress biomarkers was noted within the initial three days of exposure to continuous.
Due to continuous actibromide® dosing, mussels were unable to compensate for the reduced food intake, leading to the absence of pseudofaeces production in the treatment tanks, in contrast to observations in the control tanks. Consequently, a notable decrease in growth of mussels is inevitable (Lewis, 1985). The decline in the Condition Index (CI) of P. viridis in the actibromide® treatment tanks indicated to a reduced growth, a phenomenon previously studied in Dreissena polymorpha and Mytilus edulis regarding physiological activities (Rajagopal et al., 2003a), and in P. viridis regarding the continuous effect of low-dose chlorination (Chavan et al., 2018). One possible explanation for this phenomenon could be inadequate feeding resulting from reduced valve opening, a response previously observed in P. viridis exposed to chlorinated environments (Masilamoni et al., 2002; Rajagopal et al., 2003b).
In this study, an increase in ammonia excretion was observed in mussels treated with actibromide® concentrations of 0.2, 0.5, and 1.0 mg L− 1. Additionally, our personal observations indicated that ammonia production increases with the treatment of various concentrations of oxidizing biocide (Unpublished data). One of the reason could be due to an increase in the protein usage as a substrate during oxidative metabolism. Similar results were observed in Aulacomya ater ribbed mussels exposed to organophosphate pesticides (Führer et al., 2012) and in the bivalve Ruditapes decussates exposed to heavy metals (El-Shenawy, 2004).
The findings of this study demonstrate a significant elevation in reactive oxygen species (ROS) generation, even at the sub-lethal concentration of actibromide® (0.2 mg L− 1) tested. Furthermore, higher concentrations of actibromide® (0.5 and 1.0 mg L− 1) led to escalated levels of ROS in the digestive gland followed by gill. The previous study shows, rise in ROS production as well as increase in DNA damage in isolated tissues of Mytilus galloprovincialis (Dailianis et al., 2005) exposed to cadmium at micromolar concentration. Thermal stress, is one of the abiotic factors which directly affects organismal metabolism, leading to metabolic disorders and the accumulation of reactive oxygen species (ROS) (Bhat and Desai, 1998; Rajagopal et al., 2005). Increased ROS production in the gill tissue of P. viridis has been observed following exposure to both cold and heat stresses (Wang et al., 2018). Similarly, elevated ROS production has been observed in the gill tissue of zebrafish after cold treatment (Wu et al., 2015). Our results are in agreement with other studies that report the induction of oxidative stress after exposure to environmental pollutants and metals (Gómez-Mendikute et al., 2003; Collén et al., 2003; Lee and Shin, 2003; Koutsogiannaki et al., 2006).
ROS generates a variety of non-radicals such as superoxide radical (1O2•), hydroxyl radical (OH•) and hydrogen peroxide (H2O2•) etc. In the current study only generation of H2O2 was observed. Since high H2O2 concentration can alter vital functions in marine invertebrates (Abele-Oeschger et al., 1997), we have investigated the effect of actibromide® from sub lethal to lethal concentration. Elevated levels of H2O2 were observed in the digestive gland across varying concentrations, surpassing those detected in other assessed tissues. H2O2 can alter cell physiology through the formation of OH by Fenton reaction (Cavaletto et al., 2002). A comparable study conducted on Mytilus edulis veliger larvae, subjected to sodium hypochlorite treatment, revealed elevated levels of H2O2 upon exposure to concentrations ranging from 0.5 to 0.7 mg L− 1. Similarly, in the common sole samples, increase in H2O2 level with increase in temperature was investigated (Aslan et al., 2018).
The SOD enzyme constitutes a primary defence against oxygen toxicity by catalysing the conservation of super oxide anion to oxygen and hydrogen peroxide, which can be sequentially removed by CAT (Manduzio et al., 2004). The findings of the current study suggest that adult green mussels P. viridis exhibit SOD activity across all tissues, with higher levels observed in the digestive gland followed by gill compared to the other two tissues examined. This observation seems logical, as the digestive gland is the main source of assimilation of feed and gills are regularly exposed to high levels of oxygen due to their respiratory function (Santovito et al., 2005). Similar activity of this enzyme was reported with P. viridis with changes in seasonal variation in the reproductive cycles as well as physico-chemical parameters (Verlecar et al., 2008). This observation is also in good agreement with previous report (Filho et al., 2001). Interestingly, M. edulis exposed to chlorine exhibited elevated activity of a specific isoform of SOD, without impacting its total activity. This suggests that the differential response of isoforms should also be taken into consideration in certain cases. (Manduzio et al., 2004) for assessing toxic responses.
Organisms when experiencing stress conditions, may increase the activity of antioxidant enzymes, such as catalase (CAT), to mitigate the overproduction of reactive oxygen species (ROS), specifically higher rates of H2O2 production, and thereby prevent additional cellular damage (Regoli et al., 2014). The increase of CAT activity in the present study signifies the generation of H2O2, which also showed a strong positive correlation in the presence of actibromide®. Previous study shows that, after treatment with sub-lethal concentrations of chlorine, M. galloprovincialis showed an increase in CAT activity, which agrees with the results observed with our study (López-Galindo et al., 2010). In contrast, M. galloprovincialis exposed to metal-contaminated areas revealed inhibition of enzyme activities (Vlahogianni et al., 2007). CAT is a significant biomarker for stress induced by oxidizing agents in comparison to SOD (Vlahogianni et al., 2007). Moreover, our findings are in agreement with Verlecar et al., (2008), Chavan et al., (2018) who assessed the seasonal variation and impact of chlorine on the CAT activity in P. viridis. These results endorse the capacity of mussel cells to detoxify superoxide (O2), which is the precursor of most of the other ROS through the antioxidant enzymatic system.
GSH is a vital molecule that helps the organisms maintain their cellular health by protecting against oxidative stress and aiding in detoxification processes (Yan et al., 1997). Glutathione reductase (GR) which catalyses the reduction of oxidising glutathione (GSSG), is therefore essential for the maintenance of the GSH/GSSG ratio and the cellular redox status, protecting cells against oxidative damage (Box et al., 2007). Unlike CAT, GSH level was more in digestive gland than gill, mantle and foot in sub-lethal as well as lethal concentration of actibromide®. This result could be due to an increased consumption of glutathione required to counterbalance the stress increased due to biocide treatment. A previous study shows that, total glutathione increased slightly in Cyprinus carpio treated with sodium hypochlorite (NaClO), chlorine dioxide (ClO2) and peracetic acid (PAA) (Elia et al., 2006). In a study of New Zealand P. canaliculus mussels infected with pathogenic Vibrio sp., total glutathione levels were measured. The study found no difference in total glutathione between infected and control mussels, indicating that glutathione levels may not be affected by this particular pathogen (Alfaro and Young, 2016).
Using the comet assay we have demonstrated a dose-response relationship between the level of DNA strand breaks and the environmentally relevant concertation of actibromide®. In our present study, the mussels which were exposed to continuous mode of actibromide® showed a high % tail DNA compared to control in all the tested concentrations. Biocide addition lead to breakage of DNA strands in the mussels Dreissena polymorpha which have been used as a bioindicator of toxicity in chlorinated drinking water (Bolognesi et al., 2004) environments. Previous study on oysters Crassostrea virginica showed DNA damage in haemolymph exposed to carcinogenic spiked sediments (Nacci et al., 1996). DNA strand breaks increased after one day of exposure to benzo[a]pyrene (B[a]P) (Siua et al., 2004). Previous studies have also suggested that tail DNA content and tail moment are the most satisfactory endpoints to express the recorded DNA damage (Devaux et al., 1998; Hartmann et al., 1997). Rajagopal et al., (2003b) studied, chlorine effects on foot activity, byssus thread production, filtration activity and shell valve movement in Dreissena polymorpha, Mytilopsis leucophaeata and Mytilus edulis. Similarly, (Chavan et al., 2016) have reported genotoxic effect on green mussels P. viridis of in use levels of continuous chlorination. In the present study we used alkaline comet assay method, the most widely used method to study DNA strand break (Singh et al., 1988; Collins, 2004; Frenzilli et al., 2009).
AChE has been initially considered as a specific biomarker for organophosphates; carbamate insecticides (Galgani and Bocquene, 1989; Escartín and Porte, 1997; Canty et al., 2007) and heavy metals (Day and Scott, 1990) in the marine mollusc Mytilus edulis. The enzyme plays an important role in the functioning of neurotransmitter acetylcholinesterase to choline in cholinergic synapses and neuromuscular junctions. In this present study, AChE activity was the most responsive biomarker, showing significantly inhibition levels in haemolymph at both sub-lethal and lethal concentrations (0.2, 0.5 and 1.0 mg L− 1 TRO) of actibromide®. Studies by Vidal-Liñán et al., (2014) observed inhibition of AChE activity exposed to trace metals in gill tissue. Similar studies using the haemolymph of P. viridis showed strongly inhibition exposed to pharmaceutical drug carbamazepine (CBZ), the plasticizer bisphenol A (BAP) and the herbicide atrazine (ATZ) in a marine bivalve (Juhel et al., 2017). Previous studies reveal high AChE activity was observed in gills of Mytilus galloprovincialis and M. edulis and also in muscle tissue of fish from Mediterrian and Baltic Sea (Zinkl et al., 1987). Also, in M. galloprovincialis, changes in AChE activity were related to agricultural practices in areas where pesticides and biocides were frequently used (Escartín and Porte, 1997). Robillard et al., (2003) found lower AChE levels in the freshwater mussel Anodonta cygnea from a site in an agricultural area than in animals collected downstream from a municipal STP and at the reference site. Most studies assessing AChE levels have reported exposure to heavy metals, pesticides, pharmaceutical drugs etc (Handy et al., 2003). Till date effect of actibromide® on AChE enzyme activity of bivalves has not been investigated and the results indicates, it could serve as a sensitive biomarker of exposure to actibromide®.
The use of actibromide in heat exchangers of cooling water systems is aimed at controlling microbial growth and preventing fouling. Actibromide, a bromine-based biocide, effectively addresses issues like algae, bacteria, and slime that can impair heat exchanger performance and efficiency. By maintaining a clean and biofilm-free surface, actibromide helps ensure optimal heat transfer and prolongs the life of the equipment (Nancharaiah et a., 2000; Satyanarayanan et al., 2008). However, its use requires careful management to minimize environmental impacts, such as the potential release of bromine compounds into the water system. Proper dosing, monitoring, and adherence to environmental regulations are essential to balance effective biocide performance with minimal ecological disruption.
{kind=link}