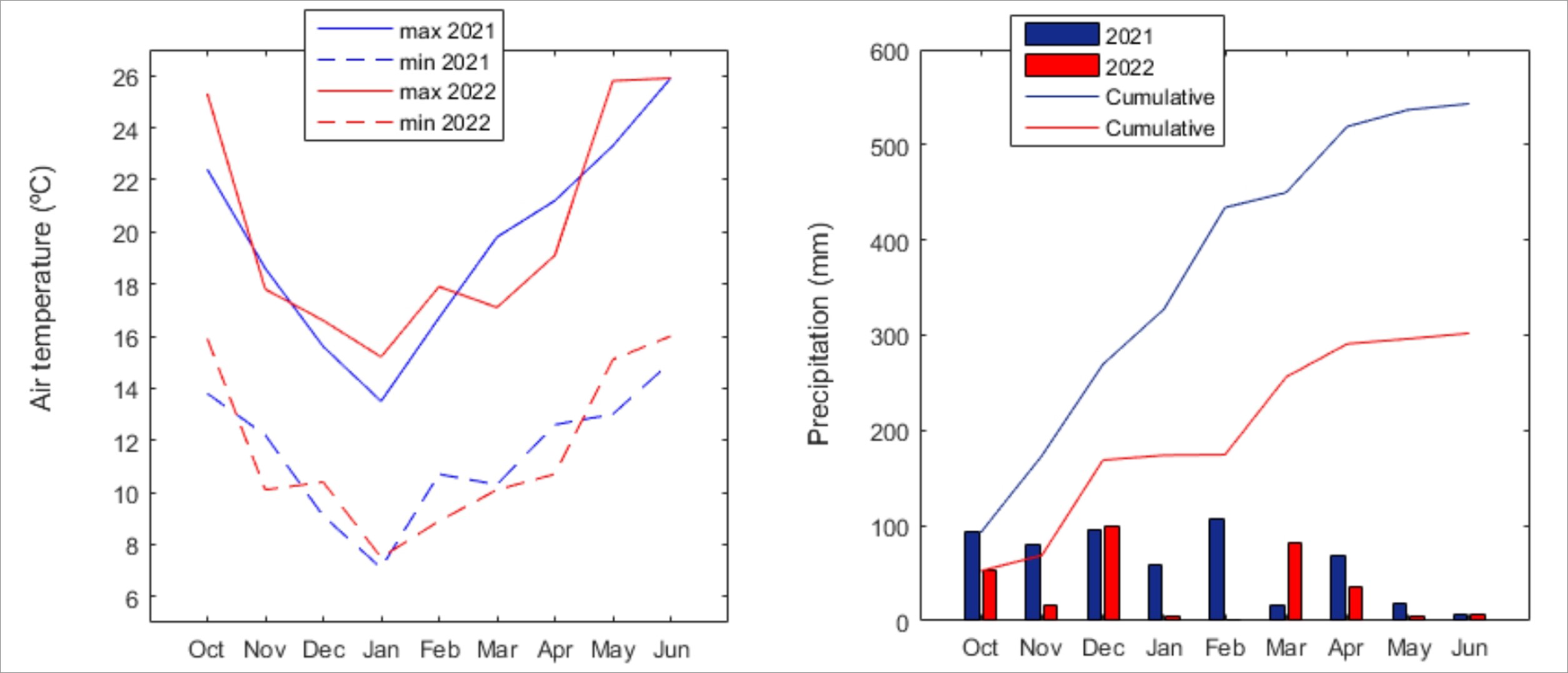
The large interannual variation in the autumn-to-spring precipitation in Portugal was evident from the contrast between 2021 and 2022, with 2021 having higher precipitation as compared to 2022 (Supplementary Fig.1). The thick mat of dry dead lawn had an average lawn shoot density of 667 shoot∙m2 and an average lawn biomass of 354 gDW∙m2. Considering all plots, a total of 17089 individuals from 46 taxa were counted and identified during the monitoring experiment. The taxa distributions among different plots were 12 taxa in E1, 22 taxa in W1 and 38 taxa in W2 (Table 1). These correspond to the β diversity, whereas the α diversity was given by the species per quadrat used in the posterior analysis.
Table 1 – Taxa observed during the monitoring and their identifier code number.
|
id. code
|
Taxa
|
E1
|
W1
|
W2
|
|
1
|
Echium plantagineum
|
x
|
|
|
|
2
|
Orobanche sp.
|
x
|
|
x
|
|
3
|
Erodium moschatum
|
x
|
x
|
|
|
4
|
Bromus sp.
|
x
|
|
|
|
5
|
Glebionis coronaria (ex Chrysantenum sp.)
|
x
|
|
x
|
|
6
|
Malva hispanica
|
x
|
|
x
|
|
7
|
Galactites tomentosus
|
x
|
x
|
x
|
|
8
|
Convulvulus sp. or Aristolochia sp.
|
x
|
x
|
x
|
|
9
|
Grass (unidentified)
|
x
|
x
|
x
|
|
10
|
Panicum repens
|
x
|
|
x
|
|
11
|
Oxalis pes-caprae
|
x
|
x
|
x
|
|
12
|
Vulpia sp.
|
|
|
x
|
|
13
|
Asparagaceae family
|
|
|
x
|
|
14
|
Stenotaphrum secundatum (Floratan)
|
|
|
x
|
|
15
|
Crepis sp.
|
|
|
x
|
|
16
|
Picris echioides
|
|
|
x
|
|
17
|
Fumaria sp.
|
|
|
x
|
|
18
|
Urtica sp.
|
|
|
x
|
|
19
|
Stellaria media
|
|
|
x
|
|
20
|
Forb (unidentified)
|
|
|
x
|
|
21
|
Polycarpon tetraphyllum
|
|
|
x
|
|
22
|
Silene sp.
|
|
|
x
|
|
23
|
Trifolium sp.
|
|
|
x
|
|
24
|
Papaver rhoeas
|
|
|
x
|
|
25
|
Sherardia arvensis
|
|
|
x
|
|
26
|
Anagallis arvensis
|
|
|
x
|
|
27
|
Arisarum simorrhinum
|
|
|
x
|
|
28
|
Solanum sp.
|
|
|
x
|
|
29
|
Calamintha nepeta
|
|
|
x
|
|
30
|
Lathyrus sp.
|
|
x
|
x
|
|
31
|
Medicago sp.
|
|
x
|
x
|
|
32
|
Conyza sp.
|
|
x
|
x
|
|
33
|
Hypochaeris glabra
|
|
x
|
x
|
|
34
|
Euphorbia sp.
|
|
x
|
x
|
|
35
|
Senecio vulgaris
|
|
x
|
x
|
|
36
|
Sonchus oleraceus
|
x
|
x
|
x
|
|
37
|
Aetheorhiza bulbosa
|
|
x
|
x
|
|
38
|
Allium sp.
|
|
x
|
x
|
|
39
|
Geranium molle
|
|
x
|
x
|
|
40
|
Andryala integrifolia
|
|
x
|
x
|
|
41
|
Gladiolus sp.
|
|
x
|
x
|
|
42
|
Urospermum picroides
|
|
x
|
|
|
43
|
unknown species (a)
|
|
x
|
|
|
44
|
unknown species (b)
|
|
x
|
|
|
45
|
Sisymbrium officinale
|
|
x
|
|
|
46
|
Centaurea sp.
|
|
x
|
|
2.1 Self-thinning and a local boundary line
Similarly to monospecific plant stands in forestry and agricultural systems, the natural biodiverse plant communities were also subject to self-thinning (Fig.2). At the beginning of the monitoring in early January, the pioneer plant communities in all plots showed high recruitment densities and low overall biomass. As time passed, the plant communities evolved to lower densities of individuals and higher stand biomass. This was the typical self-thinning trajectory that takes place during the active growth season. However, during April/May the plant communities tended to lose the larger individuals, resulting in decreased biomass. This was due to mortality of the winter-blooming individuals or taxa. From May onwards, most of the individuals and species desiccated and died.
The biodiverse communities of annual, short-lived plants that we monitored soon reached an upper boundary for the biomass-density relation (Fig.2), demonstrating that the limitations to space occupation previously found for monospecific plant stands apply as well to biodiverse plant stands. The quantile regression determined this local boundary line to be given by log10B=4.76-0.63log10D, with θ=0.566 and cosine(θ)=0.844; which places much lower than the plant IBL determined by Scrosati (2000) (Fig. 3). This local boundary line should not be considered an IBL specific of annual species since the mixed-species stands in the manipulative experiments by Black (1960), White & Harper (1970) and Bazzaz & Harper (1976) with annual species often surpassed it (Fig.3b). Furthermore, the data Weller (1989) and Scrosati (2000) used to determine the plant IBL also included annual, small-sized, short-lived species, besides perennial big trees. We question whether this local boundary represents a time-independent boundary to local communities subject to local conditions, mechanistically comparable with the universal IBL by Weller (1987) and Scrosati (2000). Alternatively, this boundary may represent the minimum self-thinning possible under the local conditions i.e., the maximum biomass increment with the minimum mortality, given by the steepest regression slope and highest intercept, and representing the community’s biomass-density timeseries under the least competition (White & Harper, 1970; Weller, 1987, 1989, 1991, Osawa & Sugita, 1989; Lonsdale & Watkinson, 1983; Lonsdale, 1990).
For unknown reasons a local boundary line was found placing substantially below the ultimate plant IBL determined by Weller (1989) and Scrosati (2000). Previously, an IBL for algae was found placing significantly above the plant IBL (Creed et al., 2019), and another IBL for seagrasses was found placing significantly below the plant IBL (Vieira et al., 2018c, 2022b). The inability to determine why any of these boundary lines place above or below the original plant IBL shows that much remains to be known about the factor(s) driving the space occupation efficiency by stands of terrestrial plants, seagrasses and seaweeds. Nevertheless, these factors affect biodiverse plant stands in the same manner that they affect mono-specific plant stands, resulting in similar dynamics, as are the case of self-thinning and a boundary line.
Competition for light has been suggested as the ultimate factor driving self-thinning and a boundary line (White & Harper, 1970; Duarte & Kalff, 1987; Morin et al., 2011; Jucker et al., 2014; Fichtner et al., 2018; Searle et al., 2022). Accordingly, Westoby (1977) proposes that self-thinning relates better with leaf area than with above-ground biomass. Our monitoring, taking place in Portugal, started in early January (peak winter), when daylength is 10h, and ended in late May, when daylength is 15h. Still, the boundary line was common to all observations within this timeframe. Therefore, we question whether, in the present case, the factor time required for self-tinning to act added to factor light. Time has been reported as a fundamental factor mediating the interactions between self-thinning and other community properties. For example, biodiversity is expected to take long to affect the productivity of plant communities (Jucker et al., 2020), whereas a meta-analysis on grazing exclusion experiments in alpine grasslands (always longer than 1 year) determined that the experiments with longer duration led to lower biomass, productivity and biodiversity (Liu et al., 2020). In the proceeding sections we show how biodiversity interacts with self-thinning and the efficiency of space occupation, and how other factors in the background, such as time and precipitation, mediate the strength of this interaction.
2.2 Effect of biodiversity (task #1)
Biodiversity influenced how close a plant community approached the local boundary line, corresponding to how efficiently it could occupy the available space. The application of indexes for species richness or evenness, in all cases showed that the maximum space occupation efficiency was attained at intermediate α diversity values (Fig.4a-d). Too few species and/or very uneven species distributions, occupied space less efficiently. This is in accordance with previous results showing that increasing biodiversity, functional diversity and structural diversity lead to forest canopies occupying space more efficiently (Pretzsch, 2014; Sapijanskas et al., 2014; Jucker et al., 2015; Williams et al., 2017; Kunz et al., 2019; Urgoiti et al., 2023; Pretzsch et al., 2024). However, there is a limitation to this effect as our study also showed that too many species and/or species distributions too even, also occupied space less efficiently. The answer to this dynamic came from a shift in paradigm. So far, biodiversity has been seen as the predictor and the efficiency of space occupation (as well as density, biomass and self-thinning) as the response. We tested the other way around, with biodiversity as a function of the efficiency of space occupation (Fig.4e,f). The α diversity attained its maximum at intermediate efficiencies of space occupation (dplant), and decreased significantly thereafter as the efficiencies of space occupation improved towards their maximum i.e., as dplant approached zero. These results suggest a feedback loop where each of biodiversity and efficiency of space occupation influences the other iteratively over time. In this feedback loop biodiversity promotes more efficient space occupations. However, as time passes, more species emerge and:
- as space occupation approaches its maximum, fierce competition eliminates the species that are less competitive (Fig.4 path pA).
- if/when the new cohorts develop fast, they may approach the boundary line without giving time and space for the emergence of additional species, thus never letting biodiversity increase substantially (Fig.4 path pB).
- the observation of the W2 community positioned substantially apart from the E1 and W1 communities in this dynamic (Fig.4a,e) indicates the presence of yet another factor acting on the background. This factor was the reduced precipitation during the winter-spring of 2022, which disabled the W2 communities to attain larger stand biomass (Fig.2). Consequently, the resulting less efficient space occupations allowed for higher biodiversity (Fig.4 path pC).
We present the results above in a manner more connected with the standard self-thinning and biomass-density graphics and theory, as well as with the general ecological theory. Maximum biodiversity was achieved at intermediate distances from the local boundary line, corresponding to intermediate space occupation efficiencies (Fig.5). Too far away from the IBL corresponded either to early colonization or to situations where the environmental conditions where too adverse for most species. On the other hand, too close to the IBL the unavailability of free space led to fierce competition eliminating the weaker individuals and species. This fits the famous ‘intermediate disturbance hypothesis’ by Connell (1978), according to which biodiversity reaches its maximum at intermediate levels of environmental disturbance and stress: where the abiotic disturbance frequency and stress are high, few species can thrive and only the fast-maturating are favoured. On the opposite extreme, where the environment is stable, and disturbance and stress are low, the biotic factors dominate the communities’ dynamics, with competitive exclusion also decreasing biodiversity (Connell, 1978; Wilson, 1990; Osman, 2015). Water availability and drought are included among the environmental stressors of terrestrial plant communities (Wilson, 1990). The wetter winter-spring to which plots E1 and W1 were subjected to allowed them to self-thin to higher stand biomass and space occupation efficiencies than plot W2, with this plot being subjected to a relatively drier winter-spring (Fig.5). Consequently, the α diversities were higher in W2 than in E1 and W1, and the β diversities were 38 taxa in W2, 22 taxa in W1 and 12 taxa in E1. Again, this fits the ‘intermediate disturbance hypothesis’ with W2 corresponding to the intermediate water stress promoting the highest biodiversity, and E1 and W1 corresponding to the low water stress where competitive exclusion eliminating the weaker species decreases the biodiversity. For the E1 and W1 plots pertaining to low environmental stress, in areas where the new annual cohorts started with already high densities and biomass - and thus, closer to the local IBL with less free space still available – self-thinning followed path pB. This path never crossed conditions for larger biodiversity. The constant unavailability of free space never allowed for the rise of biodiversity. On the other hand, in areas where the new annual cohorts started with lower densities and biomass - and thus, further away from the local IBL with more free space still available – self-thinning followed path pA. This path crossed conditions for larger biodiversity. Extending the availability of free space for longer allowed for the emergence of more species. Only when this sell-thinning path pA approached the local IBL, competition became fiercer and the weaker individuals and species were eliminated. For the W2 plot pertaining to intermediate environmental stress, the new annual cohorts started with lower densities and biomass. Self-thinning followed path pC, never closely approaching the local IBL, and thus with more free space available for the emergence of new species.
The dynamics identified and presented in this work can unify seemingly contradicting evidence and hypotheses about how biodiversity relates to stand density and productivity. The past lack of consensus and contradicting evidence may have resulted from:
(i) lack of knowledge about this dynamic that we now unveiled,
(ii) productivity achieving its maximum at roughly half (depending on the model) the environmental carrying capacity (from fundamental population dynamics), which, for plant stands, is far from the most efficient space occupations that leads to competitive exclusion.
2.3 Effect of the invasive Oxalis pes-caprae (task #2)
The emergence and replacement of Oxalis pes-caprae by autochthonous species followed well-established models of ecological succession (see Odum, 1969; Connell & Slatyer, 1977). There were two pulses of biodiversity (or cohorts of new species) (Fig.6a). The first pulse, peaking roughly around February, was in strong direct competition with Oxalis pes-caprae. During both winters (of 2021 and 2022) the invasive Oxalis pes-caprae always surged earlier than the autochthonous species. This was the core of its competitive advantage, as it occupied most of the available space, leaving little space for its late competitors (Fig.6b). This is similar to the compaction of the canopies of plants with modular construction, which became known as the Phalanx clonal growth form (Sintes et al., 2005, 2006; Brun et al., 2006a,b, 2007; Vieira et al., 2018c). In summary, during the winter bloom, Oxalis pes-caprae became the dominant competitor constraining the emergence of most other species by taking the space available ahead of the others. During January/February most of the space had already been taken (Figs 2 and 6), and in the quadrats where Oxalis had not been so efficient occupying space, the autochthonous species took it. With increasing daylength and temperatures, Oxalis pes-caprae adults were the first declining, thus opening space for other species to thrive. This second pulse, from late March onwards (Fig.6a), barely competed with the Oxalis pes-caprae decaying adults (Fig.6b). On the contrary, in the beginning of this second pulse, the new recruits benefitted from the presence of the decaying Oxalis pes-caprae adults providing protection from direct sun light and foraging birds, and reducing water losses by evaporation from soils. This was obvious during the field sampling and its signal is perceptible in Figure 6 from late March to mid-April. In this particular time-frame, higher species richness and lower dautochthonous were often associated with more Oxalis pes-caprae (i.e., dOxalis was closer to zero). Similar situations of larger adults protecting their own, as well as the community developing below them, from stress from desiccation and predation has been reported for algae (Menge, 1978; Scrosati & DeWreede, 1998; Vieira et al., 2018a,b, 2022a) seagrasses (Heck & Orth, 1980; Blundon & Kennedy, 1982; Unsworth et al., 2012; Gagnon et al., 2020; Ruiz-Montoya et al., 2021; Galván & Puente, 2023) and terrestrial plants (Gunnarsson et al., 2009; Šipoš & Kindlmann, 2013; Faralli et al., 2022). With time passing, the Oxalis pes-caprae was eradicated and the second pulse of biodiversity bursted. In summary, the decaying large adults of Oxalis pes-caprae created a microclimate more favourable for the understory recruits of autochthonous species, thus facilitating the emergence of the spring bloom. From late May onwards almost everything dried and died.
2.4 Effects of a thick mat of dead lawn (task #3)
The presence of a thick mat of dead lawn (in plot W1) did not impede the growth of new individuals and the development of two new colonization events, one during winter and another one during spring. In the comparison among the three plots, to avoid bias from eventual uneven sampling, the 15 smallest dplant, representing the 15 best space occupation efficiencies, were selected from each plot (Fig.7). A permutation test applied to this sub-set of 45 observations showed that the differences among plots were significant (p<0.0001). Post-hoc test showed that all plots differed from each other i.e., E1≠W1 (p=0.0021) W1≠W2 (p=0.0253) and E1≠W2 (p<0.0001). Thus, the developing plant community constrained by the thick mat of dead lawn during 2021 (W1) was able to grow and attain space occupation efficiencies better than those attained by the new plant community in the same plot during 2022, unconstrained by the dead lawn mat (W2). On the other hand, the community in W1 during 2021 was more biodiverse than the community in E1 during 2021, with the major difference being the presence/absence of the thick mat of dead lawn. Hence, a major effect of the thick mat of dead lawn was facilitating the colonization by new recruits, presumably by providing shelter from herbivores and creating a more favourable understory microclimate. This was similar to the effect of decaying large adults of Oxalis pes-caprae shown in the previous section 3.2. These findings refute the claim by Savory (2013a,b) that the desertification of grasslands results from the accumulation of dead plant biomass, occupying space otherwise available, and thus preventing the growth of new plants. On the contrary, the presence of dead plant biomass may act as a facilitator of the early ecological succession by creating an understory microclimate more favourable for the new recruits (our results; Odum, 1969; Connell & Slatyer, 1977). In fact, plant canopies buffer the microclimate creating more stable and favourable environments for live to thrive. This effect is evident when protecting from extreme heat or cold, and from desiccation (De Frenne, et al. 2019; Huang et al, 2024).
Besides quantitatively, the composition of the plant community was qualitatively affected by the environmental conditions. To compare the effects of the plots in the community composition, the densities (D) of each taxon were normalized: Di,norm = (Di-µD)/σD, where subscript i refers to the ith quadrat, µ is mean and σ is standard deviation. Then, their plot mean normalized densities (µW1, µE1 and µW2) were estimated and compared pair-wise (Fig.8). The names of the taxa and their identifiers are provided in Table 1. Taxa 42 to 46 were associated with plot W1, hence benefitting from the presence of the thick mat of dead loan. The dead lawn presumably created an understory micro-climate and protected from herbivory. Taxa 1 to 5 were associated to plot E1, hence benefitting from the absence of the thick mat of dead lawn and the wet winter. These taxa were competitively superior to the others in full sun and water availability. Most of the taxa 6 to 41 were associated to plot W2, hence benefitting from the absence of the thick mat of dead lawn and the dry winter. Either these taxa are competitively superior under drier conditions or, following the ‘intermediate disturbance hypothesis’ (Connell, 1978; Wilson, 1990; Osman, 2015), are generally competitively inferior but benefitted from the milder competition resulting from less efficient space occupations by the taxa that usually dominate when water abounds.
{kind=link}