Recent decades have seen significant forest expansions into treeless alpine zones across global mountain ranges, including the Alps, and this is often associated with a complex interplay of climate and land use change. The upward shift of treelines has far-reaching implications for ecosystem functioning, biodiversity, and biogeochemical cycling. However, climate variables alone explain only a fraction of treeline migration, leaving substantial research gaps regarding the influence of non-climatic factors. This study addresses these gaps by combining dendrochronological methods, high-resolution bioclimatic data, and historical land use records to examine treeline dynamics in the southern French Alps. Our results reveal a recent acceleration in tree establishment, which began in the early 2000s, and is not attributable to pastoral abandonment in the 19th century but rather to climate change. We demonstrate that historical land use changes serve as a predisposing factor for tree establishment, while recent climate change has increasingly acted as an accelerator of this dynamic. Despite identifying key climatic factors, such as thermal indicators and growing season length, that contribute to treeline shifts, our study highlights the need for further research to disentangle the specific drivers of tree recruitment and survival, particularly in the context of ongoing climate change.
Research Article
Climate, not land-use, drives a recent acceleration of Larch expansion at the forest-grassland ecotone in the Southern French Alps.
https://doi.org/10.21203/rs.3.rs-5145157/v1
This work is licensed under a CC BY 4.0 License
Version 1
posted
You are reading this latest preprint version
treeline
tree establishment wave
climate change
land use change
French Alps
Larix decidua
- A massive wave of larch establishment is detected in the southern French Alps and dated to the early 2000s using dendrochronology and high-resolution imagery.
- Tree expansion is occurring in mountain pastoral systems across various elevations, despite the absence of significant land use changes in recent decades.
- Since the late 1990s, favourable thermal and water conditions have created a climatic window of opportunity for tree establishment, with warming continuing to favour rapid expansion independent of grazing pressure.
- Although past land use changes have played a predisposing factor for larch dynamics, its spectacular acceleration over the recent decades is primarily caused by climate change.
In recent decades, forest expansions into treeless alpine areas have been documented in Asian (Cazzolla Gatti et al., 2019; Du et al., 2018), North American (Beckage et al., 2008; Trant et al., 2020) and European (Bayle et al., 2024; Carlson et al., 2014; Gehrig-Fasel et al., 2007) mountain ranges. The upward shift of the treeline ecotone significantly impacts the interaction between land and atmosphere, leading to notable changes in albedo, heat storage, evapotranspiration, and carbon sequestration (Mienna et al., 2024). It alters ecosystem biodiversity and community functioning through habitat loss for endangered species (Dirnböck et al., 2011; Pornaro et al., 2013; Rippa et al., 2011), changes in biogeochemical cycles and soil properties (Guidi et al., 2014), decline in pastoral resources (Espunyes et al., 2019) or landscape modifications (Ameztegui et al., 2021; Dirnböck et al., 2011). Given the wide array of impacts that changes in treeline elevations can have on the functioning and diversity of alpine ecosystems, it is therefore crucial to understand the underlying mechanisms of the upward shift of treelines.
As the treeline marks the physiological tolerance limit of ‘tree’ life form (Dandan et al., 2022; Körner, 2012; Körner and Paulsen, 2004), its position is partly controlled by temperature. The upward shift has thus been linked to global climate warming (Harsch et al., 2009) which is particularly pronounced in high-elevation areas (Pepin et al., 2015). However, while several review papers attribute shifting treelines primarily to global warming, climate variables account for only about 30% of global alpine treeline migration (Hansson et al., 2021; Harsch et al., 2009; Lu et al., 2021; Rees et al., 2020). This indicates that other non-climatic factors also play a significant role in mediating treeline advance (Mienna et al., 2024). Among these drivers, including topography (Ramírez et al., 2024), pedogenesis or geomorphic processes (Cudlín et al., 2017; Körner and Hoch, 2023), human land use, such as logging, anthropogenic fires, summer farming, and livestock herbivory can affect the establishment and growth of trees. In areas where land use has previously limited tree growth, current treelines are thus often legacies of historical land use which may expand towards higher elevations when anthropogenic activity reduces (Mienna et al., 2022).
In many alpine areas in Europe, the upward shift of the treeline since the mid-19th century coincides with both rising temperatures (Choler, 2023; Gobiet et al., 2014) and mass exodus leading to less intensive land use, including reduced livestock densities (Chhetri et al., 2017; Collantes, 2006; Vitali et al., 2018). Attributing advancing treelines to either of these processes remains particularly challenging, and only a few studies have attempted to quantify their respective contributions (Ameztegui et al., 2016; Gehrig-Fasel et al., 2007; Hofgaard et al., 2009; Mienna et al., 2020; Vittoz et al., 2008). In the Alps and the Pyrenees, these studies have produced contradictory results. Ameztegui et al. (2016) and Gehrig-Fasel et al. (2007) indicate that land use changes play a predominant role, with climate change influencing only a few treeline ecotones. Conversely, Vittoz et al. (2008) suggest a stronger impact of climate on these expansion dynamics. The attribution is further complicated by pulses, lags and threshold effects in the response of the treeline to environmental changes (Cullen et al., 2001; Danby and Hik, 2007; Liang and Camarero, 2018; Rupp et al., 2001). Waves in tree establishment more likely result from a combination of concurrent factors, such as specific meteorological events and abrupt land-use changes (Frei et al., 2018), than from long-term climate warming trends. Similarly, the peaks in tree establishment observed following the cessation of grazing suggest a crucial window of opportunity for trees, as biotic competition remains low before the regrowth of herbivory-sensitive plant species (Speed et al., 2010). Rupp et al. (2001) and Camarero et al. (2021) respectively explain the delayed response of treeline trees to changes by their inability to keep pace with rising temperatures, as their physiological processes are too slow.
In this context, we posit that documenting the timing of woody encroachment at a yearly resolution over extended periods, ranging from several decades to centuries, is crucial for better characterising these non-linearities and disentangling the interplay between land use legacies and climate change (Ameztegui et al., 2016; Anselmetto et al., 2024; Gehrig-Fasel et al., 2007). Schematically, two primary methods for recording tree line responses over long time periods have been used (Feuillet et al., 2020). The first considers elevational variation or forest densification using aerial photographs, which results in discrete time series of tree line positions (see, e.g., Ameztegui et al., 2016; Camarero and Gutiérrez, 2007, 2004; Gehrig‐Fasel et al., 2007; Leonelli et al., 2011). The second, based on dendrological analyses ( see, e.g., Cannone and Malfasi, 2024, for a recent review), theoretically offers continuous, annually resolved series of tree encroachment. However, the majority of the 46 studies reviewed by Canone and Malfasi (2024) group tree establishment into windows of 5 to 10 years, rather than utilising the annual resolution provided by dendrochronological analyses. These datasets are often compared with temperature series but are rarely contrasted with local, highly resolved climatic variables and detailed trajectories of land use .
To address this gap, we precisely documented larch expansion across five watersheds in the French south-western Alps using a dendrochronological approach. We employed quantile-segmented linear regressions to visualise age structures along elevation gradients and conducted breakpoint analyses to detect potential non-linearities in the spatio-temporal distribution of sampled trees. To evaluate the influence of anthropogenic factors on treeline shift, we utilised a highly resolved dataset with a finer spatio-temporal resolution than those commonly used in the scientific literature to date. This enabled us to compare larch expansion with population census records, pastoral surveys — available since the mid-19th and mid-20th centuries, respectively — and sheep stocking rates and distribution. Additionally, we considered several potential climatic drivers, rarely mentioned in previous studies, including snow melt-out date (SMOD), date of the first snowfalls (SF), heat wave index (HWI), cold wave index (CWI), and cumulative rainfall (RR), all extracted from the highly resolved climate reanalysis developed by Météo France for the French Alps and available since 1959. Specifically, we aimed to investigate (i) whether the upward expansion of larch is a continuous trend or driven by episodic establishment waves, (ii) to disentangle the respective effects of climatic and anthropogenic factors in treeline evolution and (iii) to identify the climatic drivers of the treeline shift.
2.1. Study area
We conducted this study in the upper watersheds of the Ubaye valley, located in the vicinity of the Cayolle pass in the north of the Mercantour National Park (Fig. 1). The study site spans approximately 6,500 hectares of high elevation landscapes with relatively gentle slopes, The main parental bedrock is a sandstone, locally known as the “Grès d’Annot”. Up to an elevation of around 2300 m asl, the vegetation is dominated by forests of European larches (Larix decidua Mill.), a pioneering deciduous conifer that forms monospecific stands at this elevation in this region. Above the forested areas, the landscape transitions to high elevation grasslands, sparsely vegetated areas, screes, and rocky outcrops. Scattered individual larch trees are present in these higher areas, particularly in sparsely vegetated zones. The study area experiences a continental climate characteristic of inner alpine valleys with Mediterranean influences. Ecosystems are seasonally-snow covered. This region has features of a greening hotspot, i.e. with a very significant increase of vegetation cover over the last decades (Choler & al., 2021). Recent work indicates that larch expansion is largely contributing to this pronounced greening (Bayle et al., 2024).
The landscape has been shaped by centuries of agro-sylvo-pastoralism. Summer pastures are extensively grazed by sheep herds for approximately three months, from mid-June to mid-September. The high elevation grazed lands are organised into pastoral units - i.e. mountain pastures grazed by different herds and subject to specific grazing rules. The three pastoral units in the area are Cayolle, Sanguinière and Braissette.
2.2. Tree dynamics
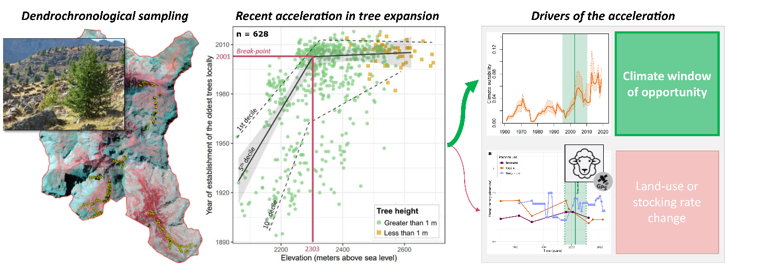
During 2022 and 2023, we conducted dendrochronological sampling campaigns across five elevation gradients ranging from 2,000 to 2,700 m asl, in areas named Braissette, Cayolle, Roche trouée, Boucharde and Eschillon (Fig. 1). These gradients extend from dense forest areas at the lower elevations to the highest tree individuals. They were selected for their varying aspects, steepness, and grazing pressures. Notably, the Roche trouée gradient has been excluded from sheep grazing since 1988. To document the colonisation patterns along these gradients, we sampled the oldest trees continuously along the elevation range. This approach allows us to determine the age structure of the larch population and trace the timeline of colonisation.
We collected cores using a Pressler increment borer, taking two cores from each tree and sapling. The first core was taken as close to the ground to precisely determine the age of the tree, while the second was taken at approximately 1 metre in height. This second sample allows us to establish a relationship between the age at ground level and at 1 metre in cases where the first core is missing. Our sampling protocol, which targeted the oldest trees in each location, enabled us to identify the onset of tree establishment (i.e. the year of initial colonisation) (Fig. 1 and 2).
Table 1. Features of the surveyed Larix decidua treeline ecotone sites in the southern French Alps.
The cores were sanded and scanned using a high-resolution scanner. Tree-rings were counted on each core using the CooRecorder software (Maxwell and Larsson, 2021). Tree-ring series were cross-dated using the Cdendro software (Maxwell and Larsson, 2021). Cross-dating was not possible for saplings due to the low number of rings. In cases where the lowest core was missing, we estimated tree age using a linear regression between the number of rings at 0 and 1 m height (Supp. Mat. 2). When increment coring failed to reach the pith, we estimated the number of missing rings using the “DistanceToPith” tool from CooRecorder (Maxwell and Larsson, 2021). This tool estimates the number of missing rings based on the radius to the estimated pith position and the spacing of previous rings. Noteworthy, our sampling protocol may slightly underestimate tree ages, as it was not always possible to core at the root collar level and, in some cases, did not reach the pith.
To visualise the age structure along the colonisation gradient, we first merged the data from the five gradients for a comprehensive analysis. We then applied the quantile-segmented linear regression method to our dataset. This method examines the relationships between different quantiles of a distribution along an explanatory variable, particularly focusing on the connections between the lower and upper quantiles and the variable. It is then particularly useful for identifying the envelope of tree age distribution along the elevation gradient. Given that the age structure appeared to change with elevation, we further refined our analysis by using segmented regressions, dividing elevation range into bins to better capture these changes. Specifically, we split elevation into 20 classes of approximately 30 m each. For each elevation class, we divided the dataset into deciles based on tree age. Finally, we performed quantile-segmented linear regressions to visualise the age structure along the elevation gradient. To conduct this analysis, we used the ‘segmented’ package in R (Muggeo, 2008).
2.3. Land use history
We sourced livestock data from pastoral surveys conducted by local authorities for three pastoral units, Braissette, Cayolle and Sanguinière. At Braissette and Cayolle, these records are partial and include the years 1950, 1963, 1972, 1995 and 2012. For the Sanguinière mountain pasture, a more extensive dataset allowed for a more detailed reconstruction of sheep numbers. For each year, we retrieved the number of sheep, the surface area of the pastoral unit and the duration of grazing on the mountain pasture. Given the changes in surface area over time, we calculated sheep density per hectare to assess variations in grazing pressure.
The number of inhabitants over the past two centuries was used as a proxy to reconstruct the history of agro-sylvo-pastoral farming. This is relevant as the workforce in traditional pastoralism was closely tied to the environmental pressure exerted by human activities (Mather et al., 1999; Rosenberg, 1988). We used population records from the four towns surrounding the study area: Uvernet-Fours, Entraunes, Saint Dalmas-le-Selvage and Allos (Source: French National Institute for Statistics and Economic Studies). These data span from the late 18th century to 2021 though data before 1850 are sparse and irregular. Subsequently, data becomes available on a much more consistent basis.
2.4. Grazing pressure
To relate grazing pressure to larch dynamics, we used individual tree location from Bayle et al. (2024) derived from photo-interpretation of very high resolution aerial infrared images. We used estimates of stocking rates obtained from position tracking of sheep (Perron Chambard et al., 2024). Briefly, 4 to 8 sheep per pastoral unit were tracked at a 10-minute frequency throughout the grazing season. From the sheep positions, three distinct behaviours were identified using Hidden Markov Models along the sheep trajectories: (i) moving; (ii) grazing and (iii) resting. The sub-trajectories for each behaviour were then converted into stocking rates through a Brownian Bridge Movement Model (Horne et al., 2007). As a result, for each pixel, we obtained a stocking rate per behaviour type. This monitoring allowed us to map stocking rate at a spatial resolution of 30 metres, expressed as the number of sheep times days spent per hectare. This protocol was applied in summers 2022 and 2023, within the Cayolle and Sanguinière pastoral units. We then merged the data from both mountain pastures and averaged it over the two years.
To assess the impact of sheep presence on trees, we set a minimum density threshold of 10 sheep/day/ha to exclude ungrazed areas and regions with minimal flock usage, where the impact of sheep was considered negligible. In this region, sheep flocks are gathered and penned overnight leading to very high local sheep densities across all three behaviours. To account for this, we applied a maximum density threshold of 250 sheep/day/ha to exclude these areas from the analysis.
We divided the space explored by the flock into two categories: (i) spaces used for moving, likely associated with high trampling rates, and (ii) spaces used for grazing and resting. For this, we analysed the pixels where the moving behaviour constituted at least 50% of the total stocking rate independently. All other pixels were used to analyse the impact of herbivory.
We used the current distribution of trees (2018) to assess the impact of sheep on tree expansion. The tree density data were aggregated on a hectare scale to better represent landscape-level dynamics and avoid very localised station effects. Finally, we compared current tree density across five quantiles of sheep stocking density, independently for both behaviours. Statistical differences between the distributions were evaluated using a Wilcoxon test, allowing for pairwise comparisons between group levels.
2.5. Climatic trends
We conducted a detailed climate analysis based on the following considerations: (i) the snow-free period is critical for tree establishment, (ii) to better understand the climatic drivers of larch expansion, it is essential to use indicators relevant to larch ecophysiology, (iii) a fine temporal resolution is necessary since seedling germination and seedling/sapling mortality are sensitive to sudden events over very short time steps. We used the S2M-reanalysis developed by Météo France for the French Alps, available since 1959 (Durand et al., 2009a, 2009b). This model integrates observed data from a network of weather stations with estimates from numerical weather forecasting models, providing hourly atmospheric data (e.g. precipitation, solar radiation) and snow metrics for 23 massifs of the French Alps across 300 m elevation bands.
Meteorological data are interpolated based on elevation, slope steepness (up to a maximum slope of 40°), slope aspect and an orographic mask with a 20km radius. The slope and mask data are extracted from a 30 m digital elevation model to account shadow effects due to terrain relief. We extracted meteorological data at our dendrochronological sampling points, corresponding to 81 pixels of 250 m each. From the S2M time series, we calculated yearly climatic indices: snow melt-out day (SMOD); snow fall day (SF); heat wave index (HWI); cold wave index (CWI); and cumulative rainfall (RR). The SMOD is defined as the first day of the year when snowpack depth falls below 10 cm, and SF is the first day late in the season when snowpack exceeds 10 cm. This threshold was chosen as it aligns with the average error of the model. These metrics allowed us to calculate the length of the snow-free period for each year, which is critical for the germination and survival of seedlings. All climate indicators were calculated during this snow-free period.
We also calculated two thermal indices: HWI and CWI, using the method of Russo et al. (2014). These indices are more informative than an average trend analysis, as they consider extreme events at a yearly resolution, which we considered the most relevant for influencing germination and seedling survival. To compute HWI, we first calculated mean daily air temperature as follows:

where Tmin and Tmax represent the minimum and maximum daily temperature, respectively.
We then computed the quantiles of Tmean for each day of the year, centred on a moving window of 15 days over the reference climate period 1991-2020. Afterward, we compared the Tmean value of each day of each year to the corresponding quantiles for the same day within the reference period. We defined a heatwave as a sequence of at least three consecutive days where Tmean exceeded the 8th decile. Finally, we computed the cumulative length of HWI per year (Fig. 5A). The definition for a cold wave is the same, but in this case, Tmean is below the 20th percentile of daily maxima.
Next, we calculated the RR (cumulative rainfall) during the snow-free period for each year (Fig. 5C).
Our objective is to integrate temperature and water-related drivers (i.e. SMOD, HWI, CWI, RR) into a function that represents the suitability of the climatic context for tree establishment (Fig. 5E). To achieve this objective, we first computed a standardised anomaly for each annual indicator relative to the 1991-2020 reference period as follows:

In this process, we applied prior knowledge about each variable. For HWI, we assumed that at this elevation, high temperatures favour tree establishment (Liu and El-Kassaby, 2015; Loranger et al., 2016). Therefore, the Max value was assigned to the longest cumulative HWI. For CWI, in high-elevation ecosystems, given that cold events are frequently associated with frost episodes, the Max value was assigned to the shortest cumulative CWI. Early snowmelt has been shown to positively affect treeline tree survival (Barbeito et al., 2012; Moir et al., 1999), so the max value for SMOD was assigned to the earliest SMOD. For RR, early tree establishment is limited by moisture availability, as drought can cause seedling damage and mortality (Loranger et al., 2016; Plesa et al., 2018). Therefore, the Max value has been assigned to the highest RR.
Next, we multiplied the standardised annual value of all the indices to construct a climate suitability function. The closer the function’s value is to 1, the more suitable the climate is for tree establishment, and conversely lower values indicate less suitable conditions. This methodology was applied independently to the 81 pixels. For each year, the median and the first and third quartiles were calculated.
The location of the climatic treeline was calculated according to the method proposed by Körner and Paulsen (2004). They define the treeline based on three criteria: (i) tree growth requires a minimum growing season length of 94 days; (ii) all days of the season must have a daily mean temperature >0.9 °C; and (iii) the mean temperature over the growing season must exceed 6.4°C. Using the air temperature provided by the S2M re-analysis, We estimated the average temperature every 25 m of elevation by applying a constant environmental lapse rate (ERL), which reflects the temperature decrease with elevation. We used an ERL of 0.56°C per 100m, consistent with the mean value observed in the Alps from late spring and summer (Rolland, 2003). Finally, we plotted the season's average temperature as a function of elevation to identify the elevation at which the 6.4°C threshold was crossed (Supp. Mat. 3). We applied an uncertainty of ± 0.05°C to this thermal threshold to determine the position of the treeline. We estimated the elevation of the climatic treeline for two distinct periods: 1961-1990 and 1991-2020.
3.1. Wave of tree establishment
In total, 628 trees were sampled across the five gradients: Braissette (n = 210), Cayolle (n = 165), Roche Trouée (n = 126), Boucharde (n = 50), and Eschillon (n = 76). The lowest sampled tree was at 2,058 m asl, and the highest at 2,693 m asl. Figure 2 shows the age structure of tree stands along elevation gradients across the five study sites. Over time, trees have progressively established at higher elevations, with younger trees appearing at higher elevations. This upward shift is corroborated by photo-interpretation showing an increase in the number of recorded trees from 312,027 in 1983 to 603,449 in 2018 (Supp. Mat. 1).
The analysis using linear regressions segmented by quantile and elevation bin, reveals a non-linear pattern in the upward movement of trees (Fig. 2). For the 5th decile curve, a simple linear regression (AIC = 534.7576, Adjusted R-squared: 0.41) was less effective in representing the data compared to a segmented regression (AIC = 499.6862, Adjusted R-squared = 0.67). Moreover, the analysis suggests at least one significant breakpoint in the relationship (P-value < 0.001), located at an elevation of approximately 2300 m a.s.l. which occurred around 2003 (Fig. 2A). The 95% uncertainty for the breakpoint year is [1995.2–2010.2]. This elevation is below both the climate treeline of the 1961–1990 period (2375 m) and the recent one of 1991–2020 (2550 m). The climate treeline has risen by 200 m over the past 30 years (Fig. 2B and Supp. Mat. 3A). Additionally, the length of the growing season has slightly increased during this period (Supp. Mat. 3B) remaining well above the 94-day threshold necessary for the transition to tree life form (Körner and Paulsen, 2004). Notably, above 2550 m asl, all larch individuals were less than 1 m in height and displayed multiple twisted stems.
3.2. No drastic land-use and stocking rate changes
An analysis of demographic trends reveals a sharp decline in population between 1840 and 1950 (Fig. 3A). The population then stabilised in the municipalities of Uvernet-Fours and Saint-Dalmas-le-Selvage,, while there was an increase in the 1960s followed by stagnation in Entraunes and Allos. The wave of tree establishment occurred during a period of demographic stability. Sheep stocking in the study area remained consistently low, with minimal variation since 1950 (Fig. 3B). Although there was a slight increase in sheep density in the 1980s, it has decreased somewhat since 1995. The stocking rate and inter-annual variations have been relatively uniform for the three pastoral units within the study area. The upper section of the Roche trouée gradient has not been grazed since 1988. In this area, the wave of tree establishment coincided with a period when the stocking rate experienced a slight decline.
The stocking rate inferred from GPS tracking of sheep shows important spatial variation (Fig. 4A). The rangeland comprises a few flat areas with night parks (in pink). Most of the pixels in the zone correspond to resting or grazing areas, with a few paths associated with sheep movements (Fig. 4A). We found no significant relationship between sheep stocking rates and tree density in 2018, and this held true for the two distinguished sheep behaviour .(Fig. 4B and Supp. Mat. 1).
3.3. Climate window of opportunity
The analysis of climate trends shows a marked reduction in the snow melt-out date (SMOD) since the 1990s, alongside a decrease in the cumulative length of cold waves and an increase in very warm episodes since 1960 (Fig. 5A and B). The trends for these thermal extremes became more pronounced during the 1990s, with the number of summer heat waves stabilising from the early 21st century onwards (Fig. 5A). Cumulative precipitation during the snow-free season shows considerable inter-annual variability and the 5-year moving average trend shows a slight decrease since the early 1990s (Fig. 5C). The probability of climate conditions being suitable for tree establishment remained low until 1995, showing only slight decadal fluctuations (Fig. 5E). However, after 1995, the probability sharply increased and peaked around 2005. It then declined slightly until 2012 before rising sharply again. The acceleration in tree establishment aligns closely with the rise in the climate suitability function (Fig. 5E). The peak observed between 1995 and 2005 (Fig. D) is synchronous with a sharp reduction in cold waves, an increase in heatwaves and an earlier snowmelt. After 2005, while the thermal indicators continue to trend towards warmer seasons, the snowpack began to melt slightly later (Fig. 5D).
In this study, we coupled tree-ring analysis with diachronic analyses of aerial photographs to precisely document treeline upward shifts since the early 20th century. This comprehensive analysis combined with a detailed record of bioclimatic indices relevant to tree establishment and historical livestock loads, allowed us to contextualise forest expansion and treeline upward shifts within a long-term framework of climate and land use changes. Our study confirmed that forest dynamics are not linear phenomena with constant tree establishment rates. Instead, tree establishment occurs episodically in response to a combination of factors. The analysis of changes in sheep stocking rates ruled out the possibility of a recent drastic change in grazing pressure as the primary driver of forest expansion Finally, our analysis supports the hypothesis that a climatic window suitable to tree germination and survival, rather than recent changes in grazing practices, has driven the observed acceleration of tree expansion.
4.1. Non-linear recolonization by larch.
The pattern of stand densification and treeline shifts, observed at our study site (Fig. 2, Fig. 6 and Supp. Mat. 1), has been widely reported across the Northern Hemisphere (Cannone and Malfasi, 2024), the European Alps (Carlson et al., 2014; Gehrig-Fasel et al., 2007), and the Pyrenees (Feuillet et al., 2020). Recently, Choler et al. (2021) identified a greening hotspot in the southern French Alps within the broader European Alps. Bayle et al. (2024) demonstrated that the expansion of larch trees largely contributed to the fast greening in this region.
Unexpectedly, our data reveal that forest expansion has not occurred at a constant rate during the 20th century (Fig. 2A). The breakpoint in tree establishment rates around the year 2000, observed simultaneously within a wide range of elevations and exposed slopes with varying exposures, is consistent with other studies that have documented a similar acceleration for Larix decidua, Pinus Cembra and Rhododendron ferrugineum in the Alps (Malfasi and Cannone, 2020; Vittoz et al., 2008) for Pinus uncinata in the Pyrenees (Batllori and Gutiérrez, 2008) and for Pinus nigra in the Apennines (Piermattei et al., 2016). Similarly, pulses in tree establishment patterns have been documented for Pinus ponderosa in Colorado (League and Veblen, 2006). Alpine studies conducted at similar elevations (above 2000 m asl) identified periods of strong acceleration, 1995–2003 (Piermattei et al., 2016) and 2000–2004 (Malfasi and Cannone, 2020) closely aligning with our findings. In the Pyrenees, Batllori and Gutiérrez (2008) observed an earlier acceleration in the 1980s.
Danby and Hik (2007) describe this pattern of episodic establishment as threshold-dependent, rather than as a gradual response of forests and treelines to climate warming. In the same vein, Camarero et al., (2021) also observed that regeneration of forest stands at various treelines coincides with periods of higher tree growth rates. However, they found that the response of tree growth to rising temperatures is non-linear and appears to be dependent on the crossing of climatic thresholds (Liang and Camarero, 2018). While long-term climate warming trends are often cited as the explanation for the waves of tree establishment, our results suggest a combination of concurrent factors, including specific meteorological events (Frei et al., 2018).
4.2. Land use changes fail to explain recent acceleration in forest expansion
The recent wave of forest expansion raises questions about the relative contributions of climate and land use changes (Ameztegui et al., 2016; Gehrig-Fasel et al., 2007). Here, we clarify these two processes by closely examining the timing of changes in land use and climate, while also exploring the factors contributing to the observed wave of tree establishment.
For this purpose, we first conducted a detailed analysis of land-use practices since World War II. Most studies focusing on the impact of land use changes on forest expansion have provided only partial analyses (Frei et al., 2023) with few undertakings in-depth historical reconstruction of stocking rates that we achieved (Anselmetto et al., 2024; Garbarino et al., 2013). Several studies have attributed forest expansion to the decline in domestic grazing over recent centuries (Gehrig-Fasel et al., 2007; Malfasi and Cannone, 2020). However, our extensive dataset of livestock timeseries covering the past 80 years shows that sheep stocking rates have remained relatively constant since the 1950s at our study site. This lack of change during the period of accelerated tree establishment challenges the hypothesis that variations in sheep density are responsible for the observed tree expansion, unless other land use practices related to grazing are overlooked (i.e., seedling/sapling removal, fire, mowing.).
Moreover, these sheep densities are low compared to those in experiments testing the effects of herbivory on the dynamics of the treeline ecotone (Speed et al., 2010). Chauchard et al., 2018) demonstrated that low livestock densities in Mediterranean mountain pastures did not prevent fir colonisation. Additionally, the observed trend of tree expansion on the Roche trouée gradient, grazed only by a small number of wild ungulates since 1988, further contradicts the idea that changes in sheep stocking are driving this process.
In parallel, we also employed a novel approach to explore potential spatial interactions between livestock presence and forest expansion (Fig. 4). By distinguishing the impacts of sheep based on their behaviour - since herbivory and trampling can have contradictory effects on woody plants (Buttler et al., 2009) - our results indicate no significant link between grazing and moving patterns and tree density (Fig. 4B). In other words, this means that the grazing and movement patterns of sheep are not influenced by the presence or absence of larch trees. This suggests that sheep presence or absence is unlikely to be the main factor driving forest expansion.
In 1993, the wolf returned to France, spreading throughout the Alps and forcing mountainous agro-pastoral systems to adapt. The main change was gathering sheep in protected night enclosures instead of allowing them to sleep freely in high-altitude, treeless areas. This shift led to more movement to these enclosures and tighter herding, which may increase trampling. However, the lack of relationship between sheep behaviour and current tree distribution suggests that a change in herding is unlikely to explain such accelerated tree expansion. Further studies are needed to clarify the impacts of livestock behaviour on woody plants.
Another possible explanation for the recent wave of tree establishment involves the biotic factors specific to larch trees. Larch cones are heavily predated by several species of fly, which decreases the number of viable seeds (Poncet et al., 2009). Larch trees therefore employ a masting strategy to maximise reproductive success (Poncet et al., 2009), resulting in highly variable seed production, with some years of very high outputs (Bisi et al., 2016). A rain of seeds could potentially explain the recent wave of tree establishment. However, larch seeds can remain dormant in the soil for several years, awaiting favourable germination conditions (Comments from Alain Roques).
Despite this, no significant forest expansion was observed until the late 1990s, suggesting that masting alone does not fully explain the dynamics we observed.
4.3. Climatic factors driving recent acceleration in treeline expansion
The abrupt increase in the probability of a climate suitable to larch establishment observed in the mid-1990s coincides closely with the wave of tree establishment at our study sites (Fig. 5). This period has seen earlier snowmelt, more frequent hot episodes, fewer cold periods, and significant cumulative rainfall. These climatic changes align with broader warming trends in the Alps (Gobiet et al., 2014), where temperature has been recognized as a critical factor for the establishment of cold ecosystem trees (Hantemirov et al., 2008; Malfasi and Cannone, 2020). Our results indicate that the combination of higher temperatures and fewer frost events, along with a reduction in snow cover duration, has created conditions favourable for tree establishment in mountainous regions. These findings are consistent with studies that have shown the positive effects of early snow melting on tree establishment (Barbeito et al., 2012; Moir et al., 1999; Vittoz et al., 2008). While early snowmelt can sometimes lead to frost damage (Havranek and Tranquillini, 1995; Neuner, 2014), the simultaneous decrease in cold episodes likely mitigate this risk in our study area. Similarly, although severe drought is driving mortality of young larch (Plesa et al., 2019, 2018), the slight downward trend in precipitation at our sites does not appear to have significantly impacted tree establishment. As our analysis focused on the snow-free period, which is crucial for seed germination and the early survival of seedlings (Germino et al., 2002; Körner, 2012), we cannot exclude the possibility that in years or locations without snow, drought and frost can still cause winter mortality of young trees (Frey, 1983).
To better understand tree establishment, we estimated a climate suitability function recognizing that tree establishment depends on a combination of multiple factors (Frei et al., 2018). Our findings suggest that the recent acceleration in tree establishment at the treeline ecotone is largely driven by climatic factors, particularly thermal indicators and the length of the growing season, which have improved the climate suitability for tree growth since the 2000s. These findings align with several studies showing that tree establishment is sensitive to climate (League and Veblen, 2006) and responds positively to rising temperatures (Malfasi and Cannone, 2020). However, there is some uncertainty regarding the exact timing of this acceleration. We cannot exclude that the climatic window may have occurred in the 1990s, a period characterised by cold, wet conditions and late snow clearance. These conditions could have led to a high germination rate in the 1990s, with favourable conditions in the subsequent decade supporting tree survival and growth. Regardless, it is clear that climate change has accelerated the upward shift of treelines.
While our study aimed at identifying relevant indicators, it did not fully assess the respective importance of various climate variables on tree establishment. Future research should therefore focus on pinpointing the specific drivers of germination and survival of treeline trees, as well as their relative significance. This could involve growing trees in communal gardens under controlled conditions or closely monitoring treeline bioclimatic conditions in the field to gain a more detailed understanding of the factors influencing tree establishment (Lechler et al., 2024).
The current climatic context still appears favourable for a high rate of tree establishment in the study area (Fig. 5). trees over 1m tall now reach elevations of 2550 m asl (i.e. the 1991–2020 treeline), and beyond that, only disturbed individuals close to the ground are found, indicating that trees are catching up with the theoretical climatic life-form limit. Furthermore, the observed rise of the climatic treeline in recent decades (an average of 200 metres) suggests that trees will continue to ascend. The acceleration in the upward shift, combined with the increasing elevation of the climatic limit, points to a potential for massive forest expansion in the southern French Alps.
However, projections show that the climate in ecosystems with seasonal snow cover will continue to warm in the coming decades (Lee et al., 2023). Camarero & Gutiérrez (2007) noted that high summer temperatures began to negatively affect Pinus uncinata establishment in the Iberian system since the 1990s. In relatively dry environments like the Southern Alps, it can therefore become the limiting factor for mountain tree expansion. The negative impact of heat episodes during dry summers on tree survival has been demonstrated in the Alps (Gazol and Camarero, 2022). Our study therefore raises questions about the future trajectory of mountain forest ecosystems.
4.4. Long-term trajectory of mountain socio-ecosystems
The elevation of the breakpoint in upward tree expansion (averaging 2300 m asl) reflects the position of the colonisation front during the early 2000s when tree establishment accelerated. This limit was below the climatic treeline of 1961–1990 and 1991–2020 periods, suggesting that the absence of trees was linked to factors other than climate (Körner, 2012). Sampling sites have been carefully selected to avoid local abiotic disturbances, such as geomorphic processes or soil deficiency, that could affect the presence of trees. Thus, we can reasonably hypothesise, following many studies have shown that most treelines in European mountains are lower than expected due to agro-sylvo-pastoral practices (Leonelli et al., 2009; Piermattei et al., 2016), that the absence of trees could be attributed to the legacy of human activities.
A comparison between historical images of the study area from 1906, which depict a treeless, barren landscape, and current images from 2014, showing tall larch forests with trees several dozen metres high, confirms not only the lowered treeline but also the significant expansion of trees over the course of the last century (Fig. 6). In this region, a massive exodus occurred from the late 18th century until shortly after World War II (Fig. 3A). This broader trend in the French Alps was largely driven by the Industrial Revolution (Collantes, 2006). The resulting labour shortages led to changes in land management practices, including the cessation of uprooting woody plants and mowing, along with the gradual abandonment of hay meadows and mountain pastures (Marini et al., 2011). As a result, maintaining mountain pastures as they were in the past became incompatible with modern farming methods.
The cessation of the practices related to the abandonment of the traditional agro-sylvo-pastoral system acts as a predisposing factor for woody plant expansion (Motta et al., 2006; Tasser et al., 2007). Trees have established themselves in areas opened by sheep movement where erosion patterns have provided regeneration niches for species like larch (Soraruf and Carrer, 2007). We consider that climate change has recently played the role of an accelerator in this ongoing forest expansion facilitating a higher rate of tree establishment at higher elevations.
The effect of changing practices, combined with climate change, has led to a complete transformation of the landscape in the study area, from open, rocky ecosystems to larch forest in just one century (Fig. 6). Such trajectories alter alpine ecosystems in different ways (Ameztegui et al., 2021): habitat loss for endangered species (Dirnböck et al., 2011; Pornaro et al., 2013; Rippa et al., 2011); changes in biogeochemical cycles and soil properties (Guidi et al., 2014); decline in pastoral resources (Espunyes et al., 2019).
Through the integration of dendrochronological methods, detailed bioclimatic analysis, and a study of the impacts of pastoralism, we have conducted a comprehensive examination of the temporality of treeline upward shifts in the southern French Alps. Our results demonstrate that tree expansion was gradual and has recently accelerated. Despite potential temporal uncertainty in the precise timing, the massive wave of tree establishment detected around the 2000s, is not due to changes in pastoral practices or sheep stocking but rather to the opening of a climatic window of opportunity.
Our study reaffirms that 19th-century land-use changes, particularly the abandonment of pastoral activities, played a crucial role in setting the stage for forest expansion. Furthermore, our findings underscore that climate change acts as a catalyst, accelerating these dynamics by creating a suitable environment for tree establishment at higher elevations.
Despite these insights, the study does not fully address the relative importance of different climatic variables influencing tree establishment. This limitation highlights the need for future research to delve deeper into the specific drivers of tree germination and survival. Ultimately, our study contributes new perspectives on the vulnerability of treeline trees during their early life stages and offers a methodological framework to better understand the effects of livestock on forest expansion.
CRediT authorship contribution statement
Baptiste Nicoud: Conceptualization, Data curation, Formal analysis, Investigation, Writing – original draft, reviewing and editing. Arthur Bayle: Conceptualization, supervision, Data curation, Writing – review and editing. Christophe Corona: Conceptualization, supervision, Data curation, Writing – review and editing. Rémy Perron-Chambard: Data curation, Writing – review and editing. Loïc Francon: Data curation, Writing – review and editing. Mathieu Fructus: Data curation. Marion Bensa: Data curation. Philippe Choler: Conceptualization, Supervision, Data curation, Writing – review and editing.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgements
This project received financial support from the ANR TOP project of the French Agence Nationale de la Recherche (ANR-20-CE32-0002). This research was conducted within the Long-Term Socio-Ecological Research (LTSER) Zone Atelier Alpes, a member of the eLTER-Europe network. LECA is part of Labex OSUG@2020 (ANR10LABX56). Thanks are also due to Marie Mathieu, Brigitte Talon and Matilda Tamalout for their help in the field as well as to Clémentine Assman, Emilie Crouzat and Laurent Dobremez for the pastoralism data provided.
- Ameztegui, A., Coll, L., Brotons, L., Ninot, J.M., 2016. Land-use legacies rather than climate change are driving the recent upward shift of the mountain tree line in the Pyrenees: Past human use drives upward tree line shift in the Pyrenees. Glob. Ecol. Biogeogr. 25, 263–273. https://doi.org/10.1111/geb.12407
- Ameztegui, A., Morán-Ordóñez, A., Márquez, A., Blázquez-Casado, Á., Pla, M., Villero, D., García, M.B., Errea, M.P., Coll, L., 2021. Forest expansion in mountain protected areas: Trends and consequences for the landscape. Landsc. Urban Plan. 216, 104240. https://doi.org/10.1016/j.landurbplan.2021.104240
- Anselmetto, N., Weisberg, P.J., Garbarino, M., 2024. Global change in the European Alps: A century of post-abandonment natural reforestation at the landscape scale. Landsc. Urban Plan. 243, 104973. https://doi.org/10.1016/j.landurbplan.2023.104973
- Barbeito, I., Dawes, M.A., Rixen, C., Senn, J., Bebi, P., 2012. Factors driving mortality and growth at treeline: a 30-year experiment of 92 000 conifers. Ecology 93, 389–401. https://doi.org/10.1890/11-0384.1
- Batllori, E., Gutiérrez, E., 2008. Regional tree line dynamics in response to global change in the Pyrenees. J. Ecol. 96, 1275–1288. https://doi.org/10.1111/j.1365-2745.2008.01429.x
- Bayle, A., Nicoud, B., Mansons, J., Francon, L., Corona, C., Choler, P., 2024. Alpine greening deciphered by forest stand and structure dynamics in advancing treelines. https://doi.org/10.21203/rs.3.rs-4668377/v1
- Beckage, B., Osborne, B., Gavin, D.G., Pucko, C., Siccama, T., Perkins, T., 2008. A rapid upward shift of a forest ecotone during 40 years of warming in the Green Mountains of Vermont. Proc. Natl. Acad. Sci. 105, 4197–4202. https://doi.org/10.1073/pnas.0708921105
- Bisi, F., von Hardenberg, J., Bertolino, S., Wauters, L.A., Imperio, S., Preatoni, D.G., Provenzale, A., Mazzamuto, M.V., Martinoli, A., 2016. Current and future conifer seed production in the Alps: testing weather factors as cues behind masting. Eur. J. For. Res. 135, 743–754. https://doi.org/10.1007/s10342-016-0969-4
- Buttler, A., Kohler, F., Gillet, F., 2009. The Swiss Mountain Wooded Pastures: Patterns and Processes, in: Rigueiro-Rodróguez, A., McAdam, J., Mosquera-Losada, M.R. (Eds.), Agroforestry in Europe: Current Status and Future Prospects, Advances in Agroforestry. Springer Netherlands, Dordrecht, pp. 377–396. https://doi.org/10.1007/978-1-4020-8272-6_19
- Camarero, J., Gutiérrez, E., 2007. Response of Pinus uncinata Recruitment to Climate Warming and Changes in Grazing Pressure in an Isolated Population of the Iberian System (NE Spain). Arct. Antarct. Alp. Res. 39, 210–217. https://doi.org/10.1657/1523-0430(2007)39[210:ROPURT]2.0.CO;2
- Camarero, J.J., Gutiérrez, E., 2004. Pace and Pattern of Recent Treeline Dynamics: Response of Ecotones to Climatic Variability in the Spanish Pyrenees. Clim. Change 63, 181–200. https://doi.org/10.1023/B:CLIM.0000018507.71343.46
- Cannone, N., Malfasi, F., 2024. Climate change triggered synchronous woody plants recruitment in the last two centuries in the treeline ecotone of the Northern Hemisphere. Sci. Total Environ. 921, 170953. https://doi.org/10.1016/j.scitotenv.2024.170953
- Carlson, B.Z., Renaud, J., Biron, P.E., Choler, P., 2014. Long-term modeling of the forest–grassland ecotone in the French Alps: implications for land management and conservation. Ecol. Appl. 24, 1213–1225. https://doi.org/10.1890/13-0910.1
- Cazzolla Gatti, R., Callaghan, T., Velichevskaya, A., Dudko, A., Fabbio, L., Battipaglia, G., Liang, J., 2019. Accelerating upward treeline shift in the Altai Mountains under last-century climate change. Sci. Rep. 9, 7678. https://doi.org/10.1038/s41598-019-44188-1
- Chauchard, S., Carcaillet, C., Guibal, F., 2018. Fir expansion not controlled by moderate densities of large herbivores: a Mediterranean mountain grassland conservation issue. Ann. For. Sci. 75, 94. https://doi.org/10.1007/s13595-018-0774-y
- Chhetri, P.K., Shrestha, K.B., Cairns, D.M., 2017. Topography and human disturbances are major controlling factors in treeline pattern at Barun and Manang area in the Nepal Himalaya. J. Mt. Sci. 14, 119–127. https://doi.org/10.1007/s11629-016-4198-6
- Choler, P., 2023. Above-treeline ecosystems facing drought: lessons from the European 2022 summer heatwave (preprint). Biodiversity and Ecosystem Function: Terrestrial. https://doi.org/10.5194/bg-2023-74
- Collantes, F., 2006. Farewell to the peasant republic: marginal rural communities and European industrialisation, 1815–1990. Agric. Hist. Rev. 54, 257–273.
- Cudlín, P., Klopčič, M., Tognetti, R., Máli&#, F., Alados, C.L., Bebi, P., Grunewald, K., Zhiyanski, M., Andonowski, V., Porta, N.L., Bratanova-Doncheva, S., Kachaunova, E., Edwards-Jonáová, M., Ninot, J.M., Rigling, A., Hofgaard, A., Hlásny, T., Skalák, P., Wielgolaski, F.E., 2017. Drivers of treeline shift in different European mountains. Clim. Res. 73, 135–150. https://doi.org/10.3354/cr01465
- Cullen, L.E., Stewart, G.H., Duncan, R.P., Palmer, J.G., 2001. Disturbance and climate warming influences on New Zealand Nothofagus tree-line population dynamics. J. Ecol. 89, 1061–1071. https://doi.org/10.1111/j.1365-2745.2001.00628.x
- Danby, R.K., Hik, D.S., 2007. Variability, contingency and rapid change in recent subarctic alpine tree line dynamics. J. Ecol. 95, 352–363. https://doi.org/10.1111/j.1365-2745.2006.01200.x
- Dandan, X., Deshuai, A., Jianqin, Z., 2022. Dynamics of the Alpine Treeline Ecotone under Global Warming: A Review. J. Resour. Ecol. 13, 476–482. https://doi.org/10.5814/j.issn.1674-764x.2022.03.012
- Dirnböck, T., Essl, F., Rabitsch, W., 2011. Disproportional risk for habitat loss of high‐altitude endemic species under climate change. Glob. Change Biol. 17, 990–996. https://doi.org/10.1111/j.1365-2486.2010.02266.x
- Du, H., Liu, J., Li, M.-H., Büntgen, U., Yang, Y., Wang, L., Wu, Z., He, H.S., 2018. Warming-induced upward migration of the alpine treeline in the Changbai Mountains, northeast China. Glob. Change Biol. 24, 1256–1266. https://doi.org/10.1111/gcb.13963
- Durand, Y., Giraud, G., Laternser, M., Etchevers, P., Mérindol, L., Lesaffre, B., 2009a. Reanalysis of 47 Years of Climate in the French Alps (1958–2005): Climatology and Trends for Snow Cover. J. Appl. Meteorol. Climatol. 48, 2487–2512. https://doi.org/10.1175/2009JAMC1810.1
- Durand, Y., Laternser, M., Giraud, G., Etchevers, P., Lesaffre, B., Mérindol, L., 2009b. Reanalysis of 44 Yr of Climate in the French Alps (1958–2002): Methodology, Model Validation, Climatology, and Trends for Air Temperature and Precipitation. J. Appl. Meteorol. Climatol. 48, 429–449. https://doi.org/10.1175/2008JAMC1808.1
- Espunyes, J., Lurgi, M., Büntgen, U., Bartolomé, J., Calleja, J.A., Gálvez‐Cerón, A., Peñuelas, J., Claramunt-López, B., Serrano, E., 2019. Different effects of alpine woody plant expansion on domestic and wild ungulates. Glob. Change Biol. 25, 1808–1819. https://doi.org/10.1111/gcb.14587
- Feuillet, T., Birre, D., Milian, J., Godard, V., Clauzel, C., Serrano-Notivoli, R., 2020. Spatial dynamics of alpine tree lines under global warming: What explains the mismatch between tree densification and elevational upward shifts at the tree line ecotone? J. Biogeogr. 47, 1056–1068. https://doi.org/10.1111/jbi.13779
- Frei, E.R., Barbeito, I., Erdle, L.M., Leibold, E., Bebi, P., 2023. Evidence for 40 Years of Treeline Shift in a Central Alpine Valley. Forests 14, 412. https://doi.org/10.3390/f14020412
- Frei, E.R., Bianchi, E., Bernareggi, G., Bebi, P., Dawes, M.A., Brown, C.D., Trant, A.J., Mamet, S.D., Rixen, C., 2018. Biotic and abiotic drivers of tree seedling recruitment across an alpine treeline ecotone. Sci. Rep. 8, 10894. https://doi.org/10.1038/s41598-018-28808-w
- Frey, W., 1983. The Influence of Snow on Growth and Survival of Planted Trees. Arct. Alp. Res. 15, 241. https://doi.org/10.2307/1550925
- Garbarino, M., Lingua, E., Weisberg, P.J., Bottero, A., Meloni, F., Motta, R., 2013. Land-use history and topographic gradients as driving factors of subalpine Larix decidua forests. Landsc. Ecol. 28, 805–817. https://doi.org/10.1007/s10980-012-9792-6
- Gazol, A., Camarero, J.J., 2022. Compound climate events increase tree drought mortality across European forests. Sci. Total Environ. 816, 151604. https://doi.org/10.1016/j.scitotenv.2021.151604
- Gehrig‐Fasel, J., Guisan, A., Zimmermann, N.E., 2007. Tree line shifts in the Swiss Alps: Climate change or land abandonment? J. Veg. Sci. 18, 571–582. https://doi.org/10.1111/j.1654-1103.2007.tb02571.x
- Germino, M.J., Smith, W.K., Resor, A.C., 2002. Conifer seedling distribution and survival in an alpine-treeline ecotone. Plant Ecol. 162, 157–168. https://doi.org/10.1023/A:1020385320738
- Gobiet, A., Kotlarski, S., Beniston, M., Heinrich, G., Rajczak, J., Stoffel, M., 2014. 21st century climate change in the European Alps—A review. Sci. Total Environ. 493, 1138–1151. https://doi.org/10.1016/j.scitotenv.2013.07.050
- Guidi, C., Magid, J., Rodeghiero, M., Gianelle, D., Vesterdal, L., 2014. Effects of forest expansion on mountain grassland: changes within soil organic carbon fractions. Plant Soil 385, 373–387. https://doi.org/10.1007/s11104-014-2315-2
- Hansson, A., Dargusch, P., Shulmeister, J., 2021. A review of modern treeline migration, the factors controlling it and the implications for carbon storage. J. Mt. Sci. 18, 291–306. https://doi.org/10.1007/s11629-020-6221-1
- Hantemirov, R.M., Surkov, A.Yu., Gorlanova, L.A., 2008. Climate changes and overstory recruitment of larch at the northern timberline in the Yamal Peninsula. Russ. J. Ecol. 39, 305–309. https://doi.org/10.1134/S1067413608050019
- Harsch, M.A., Hulme, P.E., McGlone, M.S., Duncan, R.P., 2009. Are treelines advancing? A global meta‐analysis of treeline response to climate warming. Ecol. Lett. 12, 1040–1049. https://doi.org/10.1111/j.1461-0248.2009.01355.x
- Havranek, W.M., Tranquillini, W., 1995. Physiological Processes during Winter Dormancy and Their Ecological Significance, in: Ecophysiology of Coniferous Forests. Elsevier, pp. 95–124. https://doi.org/10.1016/B978-0-08-092593-6.50010-4
- Hofgaard, A., Dalen, L., Hytteborn, H., 2009. Tree recruitment above the treeline and potential for climate-driven treeline change. J. Veg. Sci. 20, 1133–1144. https://doi.org/10.1111/j.1654-1103.2009.01114.x
- Horne, J.S., Garton, E.O., Krone, S.M., Lewis, J.S., 2007. ANALYZING ANIMAL MOVEMENTS USING BROWNIAN BRIDGES. Ecology 88, 2354–2363. https://doi.org/10.1890/06-0957.1
- Körner, C., 2021. The cold range limit of trees. Trends Ecol. Evol. 36, 979–989. https://doi.org/10.1016/j.tree.2021.06.011
- Körner, C., 2012. Alpine Treelines. Springer Basel, Basel. https://doi.org/10.1007/978-3-0348-0396-0
- Körner, C., Hoch, G., 2023. Not every high-latitude or high-elevation forest edge is a treeline. J. Biogeogr. 50, 838–845. https://doi.org/10.1111/jbi.14593
- Körner, C., Paulsen, J., 2004. A world-wide study of high altitude treeline temperatures. J. Biogeogr. 31, 713–732. https://doi.org/10.1111/j.1365-2699.2003.01043.x
- League, K., Veblen, T., 2006. Climatic variability and episodic Pinus ponderosa establishment along the forest-grassland ecotones of Colorado. For. Ecol. Manag. 228, 98–107. https://doi.org/10.1016/j.foreco.2006.02.030
- Lechler, L., Rixen, C., Bebi, P., Bavay, M., Marty, M., Barbeito, I., Dawes, M.A., Hagedorn, F., Krumm, F., Möhl, P., Schaub, M., Frei, E.R., 2024. Five decades of ecological and meteorological data enhance the mechanistic understanding of global change impacts on the treeline ecotone in the European Alps. Agric. For. Meteorol. 355, 110126. https://doi.org/10.1016/j.agrformet.2024.110126
- Lee, H., Calvin, K., Dasgupta, D., Krinner, G., Mukherji, A., Thorne, P., Trisos, C., Romero, J., Aldunce, P., Barret, K., Blanco, G., Cheung, W.W.L., Connors, S.L., Denton, F., Diongue-Niang, A., Dodman, D., Garschagen, M., Geden, O., Hayward, B., Jones, C., Jotzo, F., Krug, T., Lasco, R., Lee, Y.-Y., Masson-Delmotte, V., Meinshausen, M., Mintenbeck, K., Mokssit, A., Otto, F.E.L., Pathak, M., Pirani, A., Poloczanska, E., Pörtner, H.-O., Revi, A., Roberts, D.C., Roy, J., Ruane, A.C., Skea, J., Shukla, P.R., Slade, R., Slangen, A., Sokona, Y., Sörensson, A.A., Tignor, M., van Vuuren, D., Wei, Y.-M., Winkler, H., Zhai, P., Zommers, Z., Hourcade, J.-C., Johnson, F.X., Pachauri, S., Simpson, N.P., Singh, C., Thomas, A., Totin, E., Arias, P., Bustamante, M., Elgizouli, I., Flato, G., Howden, M., Méndez-Vallejo, C., Pereira, J.J., Pichs-Madruga, R., Rose, S.K., Saheb, Y., Sánchez Rodríguez, R., Ürge-Vorsatz, D., Xiao, C., Yassaa, N., Alegría, A., Armour, K., Bednar-Friedl, B., Blok, K., Cissé, G., Dentener, F., Eriksen, S., Fischer, E., Garner, G., Guivarch, C., Haasnoot, M., Hansen, G., Hauser, M., Hawkins, E., Hermans, T., Kopp, R., Leprince-Ringuet, N., Lewis, J., Ley, D., Ludden, C., Niamir, L., Nicholls, Z., Some, S., Szopa, S., Trewin, B., van der Wijst, K.-I., Winter, G., Witting, M., Birt, A., Ha, M., Romero, J., Kim, J., Haites, E.F., Jung, Y., Stavins, R., Birt, A., Ha, M., Orendain, D.J.A., Ignon, L., Park, S., Park, Y., 2023. IPCC, 2023: Climate Change 2023: Synthesis Report, Summary for Policymakers. Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change [Core Writing Team, H. Lee and J. Romero (eds.)]. IPCC, Geneva, Switzerland. [WWW Document]. URL https://doi.org/10.59327/IPCC/AR6-9789291691647.001 (accessed 2.26.24).
- Leonelli, G., Pelfini, M., Di Cella, U.M., 2009. Detecting Climatic Treelines in the Italian Alps: The Influence of Geomorphological Factors and Human Impacts. Phys. Geogr. 30, 338–352. https://doi.org/10.2747/0272-3646.30.4.338
- Leonelli, G., Pelfini, M., Morra di Cella, U., Garavaglia, V., 2011. Climate Warming and the Recent Treeline Shift in the European Alps: The Role of Geomorphological Factors in High-Altitude Sites. AMBIO 40, 264–273. https://doi.org/10.1007/s13280-010-0096-2
- Liang, E., Camarero, J.J., 2018. Threshold-dependent and non-linear associations between temperature and tree growth at and below the alpine treeline. Trees 32, 661–662. https://doi.org/10.1007/s00468-017-1651-2
- Liu, Y., El-Kassaby, Y.A., 2015. Timing of seed germination correlated with temperature-based environmental conditions during seed development in conifers. Seed Sci. Res. 25, 29–45. https://doi.org/10.1017/S0960258514000361
- Loranger, H., Zotz, G., Bader, M.Y., 2016. Early establishment of trees at the alpine treeline: idiosyncratic species responses to temperature-moisture interactions. AoB PLANTS 8, plw053. https://doi.org/10.1093/aobpla/plw053
- Lu, X., Camarero, J.J., Liang, E., 2021. Threshold responses of juniper tree growth and regeneration to climate warming and drought stress at alpine treeline. Trees 35, 1081–1083. https://doi.org/10.1007/s00468-021-02135-6
- Malfasi, F., Cannone, N., 2020. Climate Warming Persistence Triggered Tree Ingression After Shrub Encroachment in a High Alpine Tundra. Ecosystems 23, 1657–1675. https://doi.org/10.1007/s10021-020-00495-7
- Marini, L., Klimek, S., Battisti, A., 2011. Mitigating the impacts of the decline of traditional farming on mountain landscapes and biodiversity: a case study in the European Alps. Environ. Sci. Policy 14, 258–267. https://doi.org/10.1016/j.envsci.2010.12.003
- Mather, A.S., Fairbairn, J., Needle, C.L., 1999. The course and drivers of the forest transition: The case of France. J. Rural Stud. 15, 65–90. https://doi.org/10.1016/S0743-0167(98)00023-0
- Maxwell, R.S., Larsson, L.-A., 2021. Measuring tree-ring widths using the CooRecorder software application. Dendrochronologia 67, 125841. https://doi.org/10.1016/j.dendro.2021.125841
- Mienna, I.M., Austrheim, G., Klanderud, K., Bollandsås, O.M., Speed, J.D.M., 2022. Legacy effects of herbivory on treeline dynamics along an elevational gradient. Oecologia 198, 801–814. https://doi.org/10.1007/s00442-022-05125-8
- Mienna, I.M., Klanderud, K., Næsset, E., Gobakken, T., Bollandsås, O.M., 2024. Quantifying the roles of climate, herbivory, topography, and vegetation on tree establishment in the treeline ecotone. Ecosphere 15, e4845. https://doi.org/10.1002/ecs2.4845
- Mienna, I.M., Speed, J.D.M., Klanderud, K., Austrheim, G., Næsset, E., Bollandsås, O.M., 2020. The relative role of climate and herbivory in driving treeline dynamics along a latitudinal gradient. J. Veg. Sci. 31, 392–402. https://doi.org/10.1111/jvs.12865
- Moir, W.H., Rochelle, S.G., Schoettle, A.W., 1999. Microscale Patterns of Tree Establishment near Upper Treeline, Snowy Range, Wyoming, U.S.A. Arct. Antarct. Alp. Res. 31, 379–388. https://doi.org/10.1080/15230430.1999.12003322
- Motta, R., Morales, M., Nola, P., 2006. Human land-use, forest dynamics and tree growth at the treeline in the Western Italian Alps. Ann. For. Sci. 63, 739–747. https://doi.org/10.1051/forest:2006055
- Muggeo, V., 2008. Segmented: An R Package to Fit Regression Models With Broken-Line Relationships. R News 8, 20–25.
- Neuner, G., 2014. Frost resistance in alpine woody plants. Front. Plant Sci. 5. https://doi.org/10.3389/fpls.2014.00654
- Pepin, N., Bradley, R.S., Diaz, H.F., Baraer, M., Caceres, E.B., Forsythe, N., Fowler, H., Greenwood, G., Hashmi, M.Z., Liu, X.D., Miller, J.R., Ning, L., Ohmura, A., Palazzi, E., Rangwala, I., Schöner, W., Severskiy, I., Shahgedanova, M., Wang, M.B., Williamson, S.N., Yang, D.Q., Mountain Research Initiative EDW Working Group, 2015. Elevation-dependent warming in mountain regions of the world. Nat. Clim. Change 5, 424–430. https://doi.org/10.1038/nclimate2563
- Perron Chambard, R., Garel, M., Marchand, P., Choler, P., 2024. Fine-Scale Tracking of Sheep Grazing Pressures in Mountain Pastures: Frugal Solution and Relevant Indicators for Improved Ecosystem and Practice Management. https://doi.org/10.2139/ssrn.4875408
- Piermattei, A., Lingua, E., Urbinati, C., Garbarino, M., 2016. Pinus nigra anthropogenic treelines in the central Apennines show common pattern of tree recruitment. Eur. J. For. Res. 135, 1119–1130. https://doi.org/10.1007/s10342-016-0999-y
- Plesa, I.M., Al Hassan, M., González-Orenga, S., Sestras, A.F., Vicente, O., Prohens, J., Boscaiu, M., Sestras, R.E., 2019. Responses to Drought in Seedlings of European Larch (Larix decidua Mill.) from Several Carpathian Provenances. Forests 10, 511. https://doi.org/10.3390/f10060511
- Plesa, I.M., González-Orenga, S., Al Hassan, M., Sestras, A.F., Vicente, O., Prohens, J., Sestras, R.E., Boscaiu, M., 2018. Effects of Drought and Salinity on European Larch (Larix decidua Mill.) Seedlings. Forests 9, 320. https://doi.org/10.3390/f9060320
- Poncet, B.N., Garat, P., Manel, S., Bru, N., Sachet, J.-M., Roques, A., Despres, L., 2009. The effect of climate on masting in the European larch and on its specific seed predators. Oecologia 159, 527–537. https://doi.org/10.1007/s00442-008-1233-5
- Pornaro, C., Schneider, M.K., Macolino, S., 2013. Plant species loss due to forest succession in Alpine pastures depends on site conditions and observation scale. Biol. Conserv. 161, 213–222. https://doi.org/10.1016/j.biocon.2013.02.019
- Ramírez, L.A., Flinspach, L., Nikolić, N., Toivonen, J., Bader, M.Y., 2024. Microsite preferences of three conifers in calcareous and siliceous treeline ecotones in the French alps. Alp. Bot. https://doi.org/10.1007/s00035-024-00319-7
- Rees, W.G., Hofgaard, A., Boudreau, S., Cairns, D.M., Harper, K., Mamet, S., Mathisen, I., Swirad, Z., Tutubalina, O., 2020. Is subarctic forest advance able to keep pace with climate change? Glob. Change Biol. 26, 3965–3977. https://doi.org/10.1111/gcb.15113
- Rippa, D., Maselli, V., Soppelsa, O., Fulgione, D., 2011. The impact of agro-pastoral abandonment on the Rock Partridge Alectoris graeca in the Apennines. Ibis 153, 721–734. https://doi.org/10.1111/j.1474-919X.2011.01156.x
- Rolland, C., 2003. Spatial and Seasonal Variations of Air Temperature Lapse Rates in Alpine Regions. J. Clim. 16, 1032–1046. https://doi.org/10.1175/1520-0442(2003)016<1032:SASVOA>2.0.CO;2
- Rosenberg, H., 1988. A negotiated world : three centuries of change in a French alpine community. Toronto : University of Toronto Press.
- Rupp, T.S., Chapin, F.S., Starfield, A.M., 2001. Modeling the Influence of Topographic Barriers on Treeline Advance at the Forest-Tundra Ecotone in Northwestern Alaska.
- Soraruf, L., Carrer, M., 2007. Dynamics and structure of natural regeneration in three high elevation LTER area in the Dolomites (North-Eastern Alps, Italy). Forest@ 4. https://doi.org/10.3832/efor0448-0040177
- Speed, J.D.M., Austrheim, G., Hester, A.J., Mysterud, A., 2010. Experimental evidence for herbivore limitation of the treeline 91.
- Tasser, E., Walde, J., Tappeiner, U., Teutsch, A., Noggler, W., 2007. Land-use changes and natural reforestation in the Eastern Central Alps. Agric. Ecosyst. Environ. 118, 115–129. https://doi.org/10.1016/j.agee.2006.05.004
- Trant, A., Higgs, E., Starzomski, B.M., 2020. A century of high elevation ecosystem change in the Canadian Rocky Mountains. Sci. Rep. 10, 9698. https://doi.org/10.1038/s41598-020-66277-2
- Vitali, A., Urbinati, C., Weisberg, P.J., Urza, A.K., Garbarino, M., 2018. Effects of natural and anthropogenic drivers on land-cover change and treeline dynamics in the Apennines (Italy). J. Veg. Sci. 29, 189–199. https://doi.org/10.1111/jvs.12598
- Vittoz, P., Rulence, B., Largey, T., Freléchoux, F., 2008. Effects of Climate and Land-Use Change on the Establishment and Growth of Cembran Pine (Pinus cembra L.) over the Altitudinal Treeline Ecotone in the Central Swiss Alps. Arct. Antarct. Alp. Res. 40, 225–232. https://doi.org/10.1657/1523-0430(06-010)[VITTOZ]2.0.CO;2
Table 1 is available in the Supplementary Files section
The authors declare no competing interests.
{kind=link}