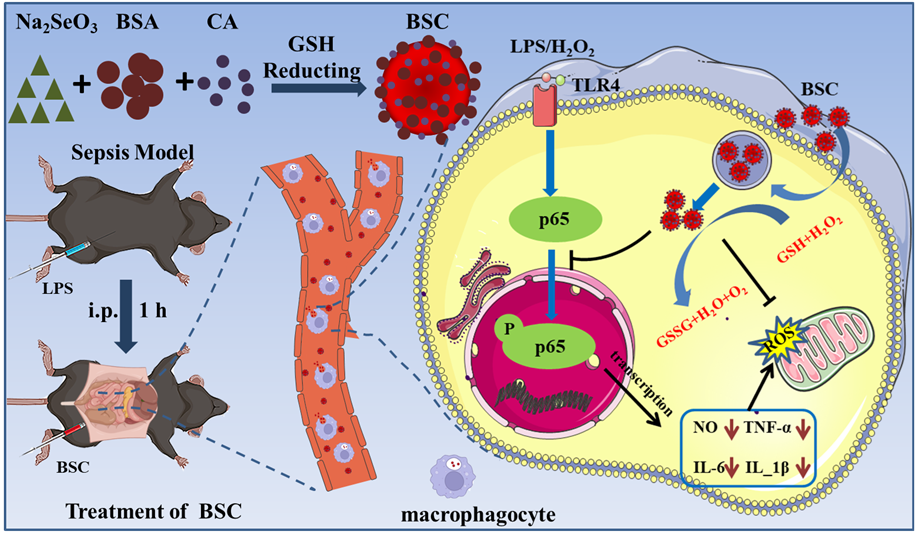
Synthesis and Characterization of BS and BSC
The BS were synthesized by one-pot method with bovine serum albumin (BSA) as a stabilizer and glutathione (GSH) as a reducing agent (Scheme 1). The transmission electron microscope (TEM) and high-resolution TEM (HRTEM) (Fig. S1) showed that the BS exhibited spheroidal morphology with an average size distribution of approximately 70 nm, which was consistent with the results obtained from Nano Particle Size and Zeta Potential Analysis (Fig. 1C and S2A). The results of Powder X-ray Diffraction (XRD) shows that the synthesized nanoparticles have no apparent lattice structure, which is consistent with the HRTEM observation. (Fig. S3) Moreover, Fourier Transform Infrared Spectroscopy (FT-IR) showed that BS appeared similar to free BSA peaks at 3282 cm− 1 (O-H), 2930 cm− 1 (C-H), 1640 cm− 1 (C-O) and 1392 cm− 1 (C-N), indicating the presence of BSA in BS and successful synthesis of BS (Fig. 1E).
The synthesis of BSC is similar to that of BS, with calycosin (CA) added to the solution before GSH reduction of sodium selenite (Na2SeO3) (Scheme 1). The synthesized BSC also appeared spheroidal morphology which is consistent with BS under TEM and HRTEM (Fig. 1A) and its size was smaller than that of BS with a size of 50 nm, which was consistent with the results of Hydrodynamic Diameter (DLS) (Fig. 1C and S2A). Similarly, XRD was employed to analyze the crystal lattice structure of the synthesized BSC, which is consistent with the BS (Fig. 1D). Zeta Potential Analysis results showed an average Zeta potential value of -30.4 mV for the synthesized BSC which is lower than that of BS alone (Fig. 1C and S2B). Energy Dispersive X-ray Spectroscopy (EDX) images revealed that selenium was the most abundant element in the synthetic nanoparticles, confirming that they were selenium nanoparticles (Fig. 1B and S4). Furthermore, XPS was employed to characterize the valence state of selenium in these particles, demonstrating that elemental selenium has a valence state of zero (Fig. 1J, S5A, and S5B). In addition to BSA-related absorption peaks in BSC in FT-IR, we observed peaks with a similar structure to CA at 1850 cm− 1 and 1020 cm− 1 which indicates the existence of CA (Fig. 1E). Furthermore, High-Performance Liquid Chromatography (HPLC) was used for quantitative analysis of CA loading. By analyzing the supernatant ultrafiltrate after centrifugation, it was found that only 50.5% of the initial amount of CA remained in solution, indicating that 49.5% of CA had been loaded onto BS (Fig. 1F). Both BS and BSC have good stability over one week without observing any polymer precipitation in H2O2, PBS and DMEM (Figure S6 and S7). Based on the fore results, we successfully fabricated the BSC.
The antioxidant mechanism of BS and BSC
Selenium, as an essential nutrient, is an important part of GPX which can catalyzes the transformation of H2O2 into H2O and O2, thus inhibiting the excessive production of ROS. In order to evaluate the GPX-like activity, we used a glutathione peroxidase assay kit to investigate their catalytic activity. We comprehensively characterized the catalytic reaction from multiple perspectives to understand the catalytic mechanism of SeNPs in exerting GPX activity. As depicted in Fig. 1G and Fig. S8, both BS and BSC exhibited GPX activity in a concentration-dependent manner. In further detecting the H2O2 scavenging efficiency of BS and BSC at 5 µg/mL was 40.31% and 40.02%, respectively ( Fig. S8 and 1G). These results demonstrated that the capability of H2O2 scavenging of BSC mainly depend on Se nanoparticles.
Many studies have been revealed the catalytic mechanism of natural GPX is the Ping-Pong mechanism. The detail catalytic process starts from the reduced state of GPX (E-SeH) is oxidized to selenite (E-SeOH) by peroxide, and then E-SeOH is reduced to the original state (E-SeH) reduced by two molecule of GSH[35]. Therefore, to further clarify the mechanism of BS and BSC mimics the catalytic activity of GPX, we explored the reaction with various methods. Firstly, supernatant at different time points and concentrations was obtained from the reaction mixture by ultrafiltration, and the selenium content in the supernatant was quantitatively analyzed by ICP-MS. Notably, with the increase in the concentration of BS and BSC, the selenium content in the supernatant increased gradually. The content of selenium element in BS supernatant increased from 0.0712 µg/mL to 0.3881 µg/mL and in BSC supernatant from 0.0727 µg/mL to 0.3771 µg/mL (Fig. 1H and S9A). Moreover, under equivalent concentrations (10 µg/mL) of BS and BSC, selenium content initially rose but then declined over time due to excess GSH re-reducing selenium ions back to elemental selenium after substrate consumption (Fig. 1I and S9B). Selenium concentrations in BS and BSC supernatants increased by 0.238 µg/mL and 0.232 µg/mL, respectively (Figs. 1I and S9B). These results indicate that the valence state of selenium may change from Se0 to SeO32−. Then the valence states of selenium in BSC before and after the reaction were further characterized by XPS analysis (Fig. 1J, 1K, and S5). The results showed that the selenite content of BSC increased, while the elemental selenium content decreased after the reaction (Fig. 1K). From the above results, we could speculate that the catalytic activity of SeNPs stems from the cyclic valence change between Se0 and SeO32−. The possible catalytic mechanism is as follows: In the first step, the low valence Se0 component was oxidized to SeO32−− by H2O2. Then, the SeO32− intermediate reacted with GSH to produce GSSG and returned to its original state (Fig. 1L). Therefore, it can be concluded that the existence of these two valences of Se components starts and maintains the catalytic reaction process of Se nanozyme.
Uptake of BS and BSC by macrophages
Cellular uptake plays a crucial role in the therapeutic effect of BSC. In this study, we studied the uptake of CA, BS, and BSC at varying concentrations in RAW264.7 cells. After co-incubation with CA, BS, and BSC, aqua regia was used to disrupt the RAW264.7, and then the selenium and CA levels in the supernatant were quantified by ICP-MS and HPLC, respectively. The results showed that the uptake of these materials was found to be concentration-dependent (Fig. 2A, 2B, and S10). The results showed that when the concentration of BS and BSC was 10 µg/mL, the content of selenium in RAW264.7 cells reached 539.1 ng/106 cells and 609.9 ng/106 cells, respectively, which indicated that selenium nanomaterials had high cellular internalization ability (Fig. 2A and S10). To visualize the uptake of nanomaterials by RAW264.7 cells, coumarin 6 (Cy6)-labeled BS and BSC were co-incubated with RAW264.7 cells, followed by observation using Inverted Fluorescence Microscopy at different time points (Fig. 2C and S11). As expected, the fluorescence intensity gradually increased within RAW264.7 cells over time, demonstrating that both materials have good cellular uptake capacity. After that, we detected the biocompatibility of CA, BS and BSC, which is an essential fore step in biological experimentation. Cytotoxicity tests showed that BS, BSC, and CA did not show significant cytotoxicity to RAW264.7 cells when the concentration of BS and BSC was 5µg/ mL− 1 and CA was 200 µM (Fig. S12A-C). In addition, the hemolytic reaction of BS and BSC did not exceed 5% at high concentrations, indicating that BS and BSC have good blood compatibility (Fig. S13). Those results indicate that BSC with good biocompatibility are suitable for treating sepsis
The antioxidant activity of BSC
Encouraged at the test-tube level, we further explored the antioxidant properties of BSC at the cellular level. To determine the effect of BSC on intracellular GPX activity, we incubated BSC with macrophages (RAW264.7 cells) for 6 hours at different concentrations and inflammatory conditions. The findings revealed a concentration-dependent relationship between GSH elimination and BS as well as BSC concentrations, and the GSH elimination rate increased with the increase of the concentration of the two compounds. indicating that BS and BSC can improve the intracellular GPX activity (Fig. 2F and S14A-C). The clearance rates of BS and BSC at 10 µg/mL were 30.27% and 32.41%, respectively (Fig. 2F and S14A-C). Furthermore, when subjected to inflammatory conditions in RAW264.7 cells activated with LPS (1 µg/mL), the GSH elimination rate was significantly higher in the groups treated with BS (27.86%) and BSC (28.30%) compared to control (2.58%), LPS (3.00%), and CA groups(0.86%) (Fig. 2G and S14D). Hence, our results demonstrate that both BS and BSC can effectively enhance intracellular GPX activity.
Based on this premise, we conducted further investigations into the impact of BSC in intracellular ROS. As shown in Fig. 2E, we used DCFH-DA to detect the total intracellular ROS level. The analysis results showed that BSC had the strongest effect and could effectively reduce the intracellular ROS level. Then we employed a Fluorescent Microplate Reader to quantify intracellular ROS levels. After normalization, the fluorescence intensity of LPS group, CA group, BS group and BSC group were 1.86, 1.79, 1.46 and 1.34, respectively, and the fluorescence intensity in RAW264.7 cells activated with LPS (1 µg/mL) increased compared with untreated cells, but after treatment with CA, BS, and BSC, the fluorescence was decreased. And BSC showed a higher activity, which exhibits the synergistic antioxidant properties of BS and CA (Fig. 2D).
Subsequently, we investigated the protective effect of BSC against ROS-induced oxidative damage in normal cells. H2O2 is a pivotal component in ROS production due to its stability and ability to diffuse across cell membranes. Therefore, H2O2 was employed as an inducer to provoke damage in RAW264.7 and HUVEC cells, enabling us to assess the protective effects of CA, BS, and BSC. The results demonstrated that CA, BS, and BSC exhibited significant protective effects on both RAW264.7 cells and HUVEC cells (Fig. 2H and 2I). After calculation, Compared with the LPS group (66.33%), the cell viability of RAW264.7 cells treated with CA, BS, and BSC was restored to 75.05%, 80.89%, and 95.45%, respectively (Fig. 2H). Similarly, compared with the LPS group (51.58%), the cell viability of HUVEC cells in the CA group, BS group, and BSC group recovered to 67.19%, 77.19%, and 85.71%, respectively (Fig. 2I). Notably, The protective effect of BSC is higher than that of CA and BS alone, suggesting that BSC have a synergistic protective mechanism. Therefore, it indicated that BSC may be an excellent candidate to protect cells against ROS damage.
The anti-inflammatory activity of BSC
Uncontrollable ROS and sustained oxidative stress will induce irreversible inflammatory injury. Moreover, it initiates the circulatory process that inflammatory response and ROS can promote each other, amplifying oxidative stress throughout the body until it develops into sepsis of systemic inflammation. Therefore, scavenging ROS is an effective approach to inhibit inflammatory signaling. In addition to effectively clearing excess ROS, BS and CA have also been reported to be able to clear inflammation-related cytokines, including NO, IL-6, IL-1β, and TNF-α. Then, we further detected the inflammatory response in macrophages after being treated with CA, BS, and BSC, respectively. The results of our study on the inhibition of NO by different concentrations of BS and CA demonstrated that BS could effectively inhibit the release of NO even at low concentrations (1.25 µM). Both BS and CA exhibited a concentration-dependent inhibition effect on NO production, with higher concentrations resulting in decreased levels of NO (Fig. 3A and 3B). Additionally, we investigated the inhibitory effects of BSC on NO. Compared to the Control group, LPS group, CA group, and BS group, it was observed that BSC exerted a superior inhibitory effect on NO compared to either BS or CA alone (Fig. 3C). The release of NO decreased from 2.78 times (the LPS group) to 1.56 times (the BSC group) (Fig. 3C). Furthermore, as shown in Fig. 3D-F, after activating (the LPS group), the levels of three pro-inflammatory cytokines (TNF-α, IL-6, and IL-1β) increased. However, after treatment with CA, BS, and BSC, the levels of pro-inflammatory cytokines were decreased. Notably, BSC displayed superior inhibitory efficacy compared to both BS and CA. However, the synergistic inhibition of TNF-α, IL-6, and IL-1β by BSC is not obvious, which may be due to the difficulty of sustained release of CA at the cellular level. Western blot demonstrated that different experimental conditions did not impact the expression of p65 (Fig. 3G and 3I). However, they did influence the phosphorylation status of the p65 protein(Fig. 3G and 3H). Remarkably, through synergistic inhibition of p65 phosphorylation, BSC effectively mitigated the inflammatory response in RAW264.7 cells.
The therapeutic effect of BSC on mice with sepsis
To further investigate the therapeutic effect of BSC on sepsis, we first studied the toxicity of BSC in healthy mice. After injecting 2 mg/Kg and 4 mg/Kg of BSC into healthy mice, we found that none of the mice died and no significant change in body weight occurred within 15 days. (Fig. S16A and S16B). Blood routine analysis after 15 days showed that BSC did not show obvious toxic side effects on blood cells (Fig. S16C). Encouraged by the above results, then we observed the therapeutic effect of BSC in a well-established mouse model of LPS-induced endotoxemia (Fig. 4A). Mice that received LPS via intraperitoneal (i.p.) administration (7.5 mg/kg) all died within three days. At a dose of 0.5 mg/kg, CA did not enhance the 15-day survival rate of septic mice. Conversely, the survival rates of both BS and BSC were significantly enhanced to 25% and 60%, respectively (Fig. 4B). Although notable body weight loss was initially observed, the mice treated with the BSC all started to recover after day 3 (Fig. 4D).
To gain insights into the distribution and metabolism of the nanomaterial BSC in sepsis mice, we employed 177Lu for surface labeling of BSC[36]. Within the initial 2 hours, the nanomaterials were extensively distributed throughout the abdominal cavity, with the discernible presence of BSC in vital organs such as the liver, spleen, lung, kidney, and gastrointestinal tract. Subsequently, over time, circulating 177Lu-labeled BSC were also detected in brain tissue along with cardiac and select joint tissues (Fig. 4C). In summary, it is clear that metabolic clearance of BSC primarily occurs through fecal and urinary excretion pathways while respiratory elimination is also observed.
In addition, the serum GPX activity of sepsis mice increased four hours after BSC administration. The findings revealed no significant disparity in GPX enzyme activity among the Control, LPS, and CA group. Conversely, a notable elevation in GPX activity was observed in septic mice treated with BS and BSC, increasing by 34.80% and 31.28%, respectively (Fig. 4E and S15). This augmentation of GPX activity in septic mice holds potential benefits for mitigating oxidative stress levels and subsequently controlling the progression of sepsis.
To further evaluate the therapeutic efficacy of the BSC, cytokine levels in the plasma of LPS-treated mice were tested by enzyme-linked immunosorbent assay (ELISA). The concentration of pro-inflammatory cytokines IL-6, IL-1β, and TNF-α, the key factors responsible for the cytokine storm and the high mortality in sepsis mice, were decreased with various degree after BSC administration (Fig. 4F, 4G, and 4H). Compared with the control group, the levels of IL-6, IL-1β and TNF-α in serum of LPS-induced sepsis mice were 11.10 times, 9.54 times and 10.30 times of normal levels, respectively. After treatment with BSC, IL-6, IL-1β and TNF-α were reduced to 2.15, 2.56 and 6.38 times of normal levels, respectively. The expression of multiple pro-inflammatory was decreased, suggesting an improved immune status in these mice. In addition, BSC showed obvious synergistic inhibition on the production of IL-6, IL-1β and TNF-α, which may be due to the continuous decomposition of nanomaterials and the continuous release of CA.
Sepsis leads to tissue and organ injuries that can harm the body's overall health. For instance, lung injury resulting from sepsis is often indicated by significantly elevated levels of alanine aminotransferase (ALT) and aspartate aminotransferase (AST) in the bloodstream. Therefore, measuring ALT and AST concentrations in serum allows for assessing the extent of lung injury. We further evaluated routine blood and blood biochemical indicators in mice. Our findings reveal that 24 hours after LPS injection-induced sepsis, mice exhibited significantly increased levels of AST and ALT along with lung injury symptoms; however, upon administration of BSC post-injection, both AST and ALT levels were markedly reduced, indicating that BSC can reduce lung injury caused by sepsis (Fig. 5A and 5B). The AST level decreased from 156U/L to 85U/L, and the ALT level from 494U/L to 249U/L (Fig. 5A and 5B). Furthermore, following LPS injection-induced sepsis in mice models, there was a sharp decrease observed in immune cell counts, including white blood cells (WBC), lymphocytes (Lym), monocytes (Mon), and eosinophils (Eso). However, BSC treatment prevented WBC and Lym depletion compared to other treatment groups, and the number of WBC and Lym cells remained at 44.04% and 53.07%, respectively (Fig. 5C and 5D). Moreover, BSC treatment resulted in an 7.52-fold and 0.93-fold increase in Mon and Eso numbers, respectively, compared to the control group, which is beneficial for combating foreign pathogens while preventing immunosuppression compared to the control group (Fig. 5E and 5F).
The sepsis can cause systemic inflammation which severely damage lead to organic failure and death. Therefore, therapy the organs dysfunction inducing by sepsis is urgent. we conducted H&E staining and Tunnel staining analysis on major organs such as the liver, spleen, and kidney from different treatment groups to evaluate the protective effect of BSC. The results showed that the necrosis of liver and kidney cells was significantly reduced after BSC treatment. In addition, the lymphocyte infiltration of liver and kidney tissues and the apoptosis of spleen lymphocytes were also inhibited (Fig. 5G). In short, these comprehensive results confirmed that BSC have an excellent therapeutic effect on sepsis, indicating that it has potential biomedical application value.
{kind=link}