Species identification of field samples
Species identification was performed on a total of 441 individual mosquitoes collected in Tiefora, Burkina Faso and Bahirdar, Ethiopia, consisting of 248 and 107 samples respectively. The Burkina Faso collections consisted of 134 (54%) An. arabiensis, 21 (8.5%) An. coluzzii, 92 (38.1%) An. gambiae and 1 (0.4%) An. gambiae x An. coluzzii hybrid. In contrast, the Bahirdar samples were all An. arabiensis (107 samples). Table 1 shows the number of control unexposed mosquitoes and those alive and dead following pyrethroid exposure.
Table 1
Breakdown of the mosquito samples by species, mortality status and country. Table 1 shows the country and species for control unexposed mosquitoes, alive, dead and total and those that were exposed to 5XDD Deltamethrin through WHO tube bioassay, alive, dead and total. The final column shows total number of mosquitoes tested for each row. The final row shows the total across both countries.
| Country/Species | Control | Control Total | Exposed | Exposed Total | Total |
| Alive | Dead | Alive | Dead | |
| Burkina Faso | 58 | | 58 | 88 | 102 | 190 | 248 |
| An. arabiensis | 48 | | 48 | 18 | 68 | 86 | 134 |
| An. coluzzii | 4 | | 4 | 12 | 5 | 17 | 21 |
| An. gambiae | 6 | | 6 | 57 | 29 | 86 | 92 |
| Hybrid (An. gambiae x An. coluzzii) | | | | 1 | | 1 | 1 |
| Ethiopia | 32 | 2 | 34 | 15 | 58 | 73 | 107 |
| An. arabiensis | 32 | 2 | 34 | 15 | 58 | 73 | 107 |
| Grand Total | 90 | 2 | 92 | 103 | 160 | 263 | 355 |
Resistance levels
Resistance to the pyrethroid deltamethrin (Fig. 1A) was highest in field-caught An. gambiae with 66.3% of the mosquitoes surviving 5XDD exposure, in contrast to 20.9% survivorship of An. arabiensis. An. coluzzii from Burkina Faso also showed similarly high survivorship (70.6%) but with only 17 samples. An. arabiensis samples from Ethiopia showed similar resistance levels to An. arabiensis in Burkina Faso with 20.5% surviving 5XDD. The lab colonies were exposed to 1XDD of deltamethrin, with 41% An. arabiensis surviving and 67.3% of An. coluzzii. Each field-caught individual used for subsequent microbiome work was then assessed for the presence of kdr-L995F and ACE-1 mutations (Fig. 1B); kdr-L995F was present in 42.6% of the An. arabiensis from Ethiopia (29.4% in dead and 33.3% in alive) and ACE-1 at 6.4% (3% in dead and 10% alive). In Burkina Faso, kdr-L995F was present at 54.4% in An. arabiensis (66.7% alive and 36.7% dead), and was significantly associated with survival (pχ2 = 0.0379); An. coluzzii at 60% (75% alive and 30% dead, pχ2 = 0.045) and An. gambiae at 62.5% frequency (63.3% alive and 61.9% dead, pχ2 = 0.88). L995S was not tested here, although is likely present in An. arabiensis from Burkina Faso [36]. ACE-1 was at low frequency across all Burkina samples, with 4.4% in An. arabiensis, 3.3% in An. coluzzii and 19.5% in An. gambiae.
Microbiome diversity
16S sequencing was then performed on alive and dead mosquitoes from pools of each species from each location. No differences in alpha diversity were observed, indicating no difference in operational taxonomic unit (OTU) richness; however, beta diversity was significantly different between the countries and the interaction term of countries and alive/dead status (PANOVA = 0.001 and 0.008 respectively) signifying diversity between these factors. No differences were observed for beta dispersion across the countries, demonstrating similar variances between groups. A Bray-Curtis multidimensional scaling plot (Fig. 2A) shows that the samples largely separate on location, with some notable exceptions. Several samples from the An. arabiensis colony and one sample from the An. coluzzii colony overlap with the Burkina Faso samples, whilst one An. arabiensis pool from Burkina Faso overlaps with those from the An. arabiensis colony; this may indicate some conservation of original microbiome stochastically in individuals across many generations and across multiple localities. There is no clear distinction between the samples from Burkina Faso based on species; this lack of separation might be expected, as the individuals, despite originating from different larval environments, were later pooled. Indeed, sharing a niche during the aquatic stage has been shown to significantly influence microbial composition [37]. Interestingly, whilst the samples from Burkina Faso and the colonies show no separation of those mosquitoes surviving or dying after insecticide exposure, there is clear separation of the Ethiopian samples indicating that microbiome composition is linked to insecticide resistance in these An. arabiensis samples. These samples were collected from the same collection site, potentially explaining the clearer assoaciation.
The abundance plots (Fig. 2B) show 2–6 highly abundant genera across each population, An. arabiensis from Ethiopia are dominated by Erwiniaceae, which align to an unspecified taxon, Salmonella and Pantoea. Six OTUs are above 10% abundance in the colonised An. coluzzii including Asaia, Salmonella, an unspecified Erwiniaceae, Swaminathania, Elizabethkingia anophelis and Enterobacter amnigena whilst An. coluzzii from Burkina Faso are dominated by Swaminathania, with Asaia and Salmonella having > 5% abundance. Similarly, An. gambiae from Burkina have these three OTUs over 5% abundance but uniquely have Tanticharoenia. An. arabiensis from Burkina Faso again shows similar dominant taxon, overlapping entirely with the other species. The An. arabiensis colony is dominated by two taxa: Elizabethkingia anophelis and an unspecified Swaminathania.
Bacteria associated with survival to pyrethroid exposure
To explore any association with insecticide resistance, the pools were split by country and association with survival assessed for OTUs representing > 1% of the overall abundance. Field caught samples from Burkina Faso had one genus associated with survival, Sphingomonas, which was at significantly higher abundance in survivors (pANOVA = 0.029). Similarly, the An. arabiensis colony had a low abundant unspecified gammaproteobacteria which was at higher abundance in dead mosquitoes (pANOVA = 0.03). Unlike these tenuous associations, the field caught An. arabiensis from Ethiopia had clear associations with bacterial genera showing significant association with survivorship. As with Burkina Faso, Sphingomonas was significantly associated with exposure (pANOVA = 0.045); however, it was found at higher abundance in dead mosquitoes. Perhaps the most interesting is a highly significant association of Pantoea with survival (pANOVA = 0.0067) at > 12% abundance in alive mosquitoes compared to ~ 2% in dead mosquitoes. Similarly, the unspecified Erwiniaceae is at ~ 66% abundance in alive mosquitoes and 12% in dead mosquitoes (pANOVA = 0.0082), whilst Salmonella is at ~ 14% in alive mosquitoes and 47% in dead mosquitoes, although this is not significant due to high variation (pANOVA = 0.11) (Fig. 3). The other significant genera at significant higher abundance in survivors include Escherichia and Kosakonia, whilst Rhizobium and Methylbacterium-Methylorbrum are at higher abundance in dead mosquitoes; however, the abundance of these bacteria is on average < 1%.
Characterisation of insecticide resistance in An. arabiensis from Ethiopia
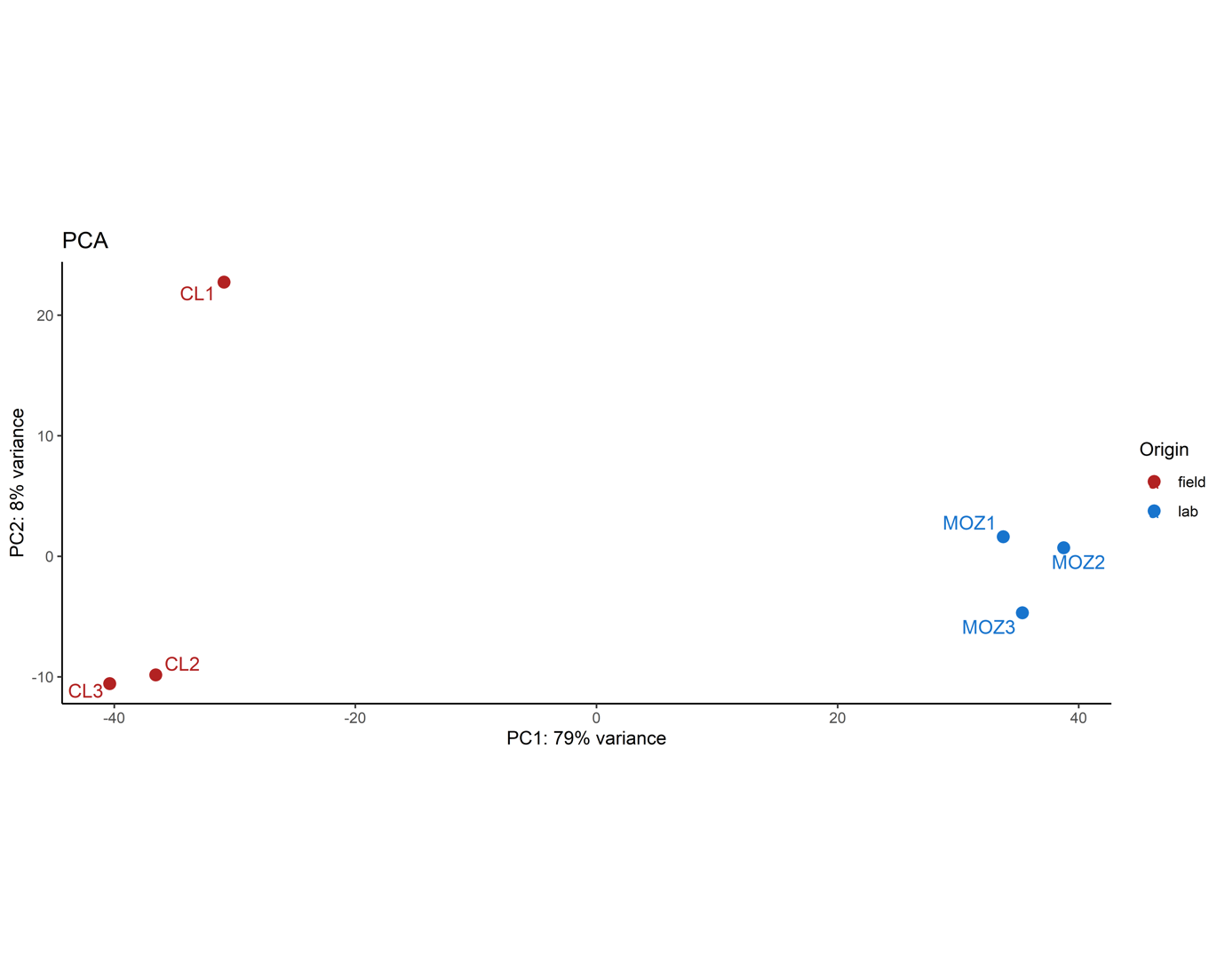
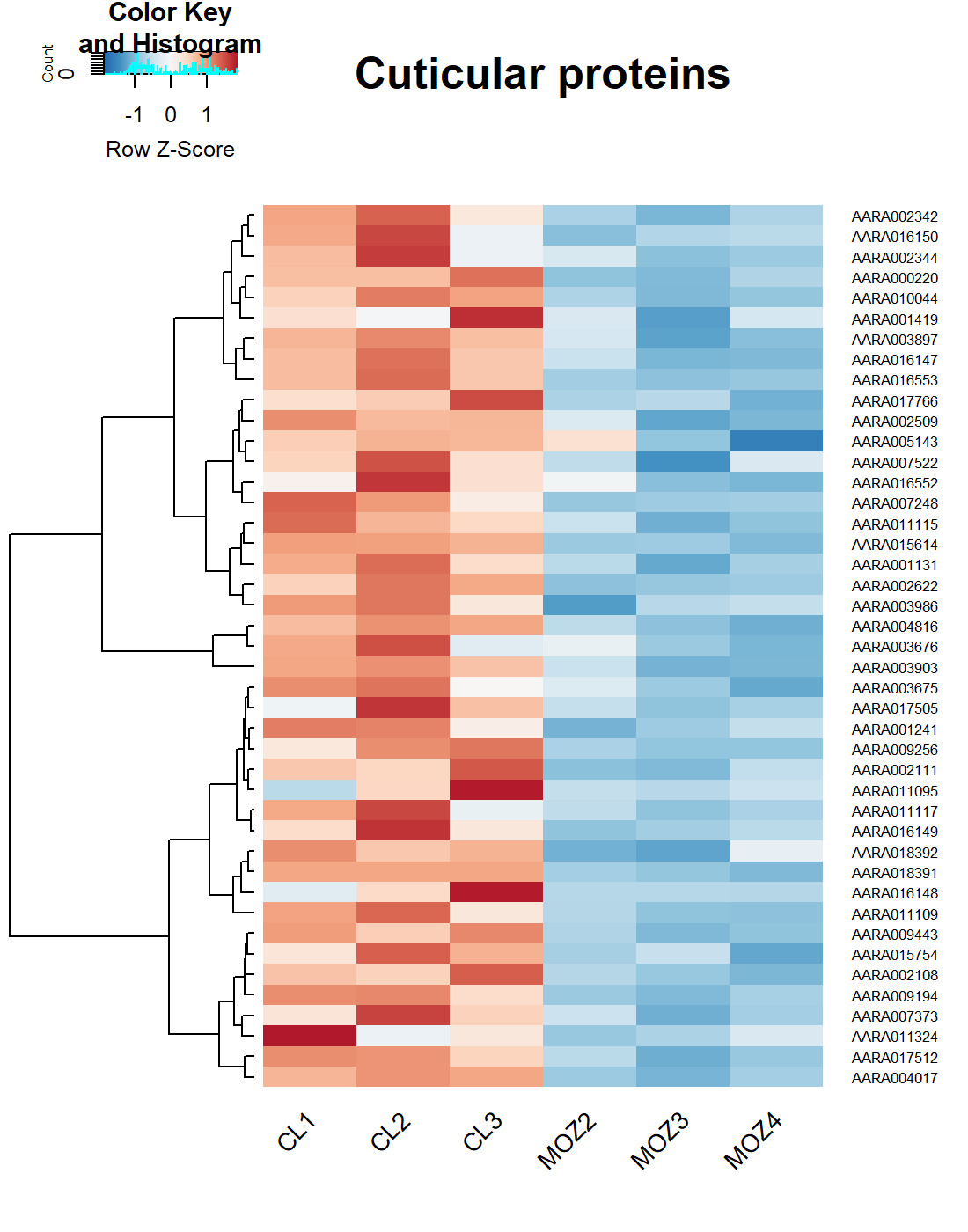
To determine the mechanisms of genetically-driven pyrethroid resistance in the field caught An. arabiensis from Ethiopia, RNAseq was performed. PCA demonstrated separation of field mosquitoes from Bahirdar and the lab-susceptible Moz (Supplementary Fig. 1). In total, 2666 genes (26.3%) were significantly overexpressed and 2958 (29.1%) were significantly down-regulated (Fig. 4A, Supplementary Table 1). GO terms show significant enrichment numerous terms, including cellular respiration (p = 6.07e-7), ATP synthase (p = 6.15e-3), oxidoreduction activity (p = 2.09e-5), ion transport (p = 1.31e-9) and synapse (p = 1.21e-3). Four MetaCyc pathways showed significant enrichment, including lipoprotein post-translational modification (p = 1.25e-5), aerobic respiration (p = 4.07e-3), Fe(II) oxidation (p = 6e-3) and plasmalogen biosynthesis (p = 1.47e-2) (Supplementary Table 2). Of the detoxification genes, 23 ABC transporters, 33 cytochrome P450s, 10 GSTs, 5 COEs and 2 UGTs are overexpressed (Figs. 4B-C). These genes include CYP9K1, CYP6P4, CYP6AA1, CYP6Z3 and CYP6M2, which are all known pyrethroid metabolisers in the An. gambiae complex [18, 19, 38, 39] and CYP4G16 linked to cuticular thickening [40]. Additionally, the syntenic ortholog of ABCH2 [41], 3 CSPs [25], 3 alpha-crystallins, 3 hexamerins [14] and 24 cuticular genes are overexpressed (Supplementary Fig. 2). AARA016988 is the homolog of the hexamerin AGAP001345 [14] and is the second most highly significantly over-expressed gene at 175x, whilst AARA016998, the homolog of SAP2 [25], is the fourth with a fold change of 18.45. ABCA7 and ABCG2 homologs are present in the top 20 significantly overexpressed genes, whilst 5 of the top 20 are serine proteases.
GO term enrichments of significantly down regulated genes include nucleic acid binding (p = 2.19e-17), gene expression (p = 8.55e-31), cellular response to stress (p = 5.17e-13) and ribosome biogenesis (p = 8.07e-4). The KEGG pathway related to biotin metabolism (p = 1.88e-3) is also significantly enriched (Supplementary Table 2). Taken together, these indicate large metabolic changes between these populations.
Comparison of An. arabiensis from Ethiopia and Burkina Faso
A prior dataset of permethrin resistant An. arabiensis from Asendabo, approximately 425km further south [42] was compared with the results from Bahirdar. 1443 genes were significant across both sites, 607 are consistently up-regulated and 412 consistently down regulated (Supplementary Table 3). Consistently up-regulated genes include CYP6P4, CYP6M2, CYP6P3, CYP9J5 and CYP9K1, all previously implicated in pyrethroid resistance [38, 39, 43, 44]. Furthermore, GSTE2, GSTE7, GSTD3 a chemosensory protein homolog and two ABCG transcripts are included in this list. Enrichments include oxidoreduction-driven active transmembrane transporter activity (p = 2.09e-17), oxidoreductase activity (p = 1.61e-12), electron transfer activity (p = 3.82e-16), cellular respiration (p = 1.82-20) and mitochondrion (p = 2.89e-11). Consistently down regulated genes include response to stress (p = 1.61e-2).
Similarly, a previously published data is available for An. arabiensis from Gaoua, located in southwest Burkina Faso [36] and this was compared to the Ethiopian samples (Supplementary Table 4). A pairwise comparison with the samples from Bahirdar shows 2762 genes significant across both sites, of these 209 are commonly up-regulated and 176 are commonly down regulated. Consistently up-regulated genes include CYP6AA2, CYP6AG2, CYP6AK1, CYP6P1 and CYP6P4. Of the other detoxification family members, ABCA2, GSTE7, GSTS1, GSTMS3 and a UGT (AARA006222). Enriched GO terms relate to transporter activity (p = 4.95e-2).
A three-way comparison shows 726 genes commonly differentially expressed, of which 55 are commonly up-regulated and 22 genes that are commonly down-regulated (Supplementary Table 5). GSTE7, CYP6AK1, CYP6P4, a UGT (AARA006222), one COE (AARA016325) and multiple serine protease-related proteins are commonly over expressed. The consistent overexpression in these vastly different populations indicates a key role in insecticide resistance in these populations.
{kind=link}
{kind=link}