The primary findings of the present study were that SB modulated the gut microbiota profile of T2D mice and elevated the fecal concentration of butyrate, which alleviated stroke outcome independent from a T2D background. This protective effect was related to the enhancement of BBB integrity and preservation of the endothelial glycocalyx in the ischemic brain.
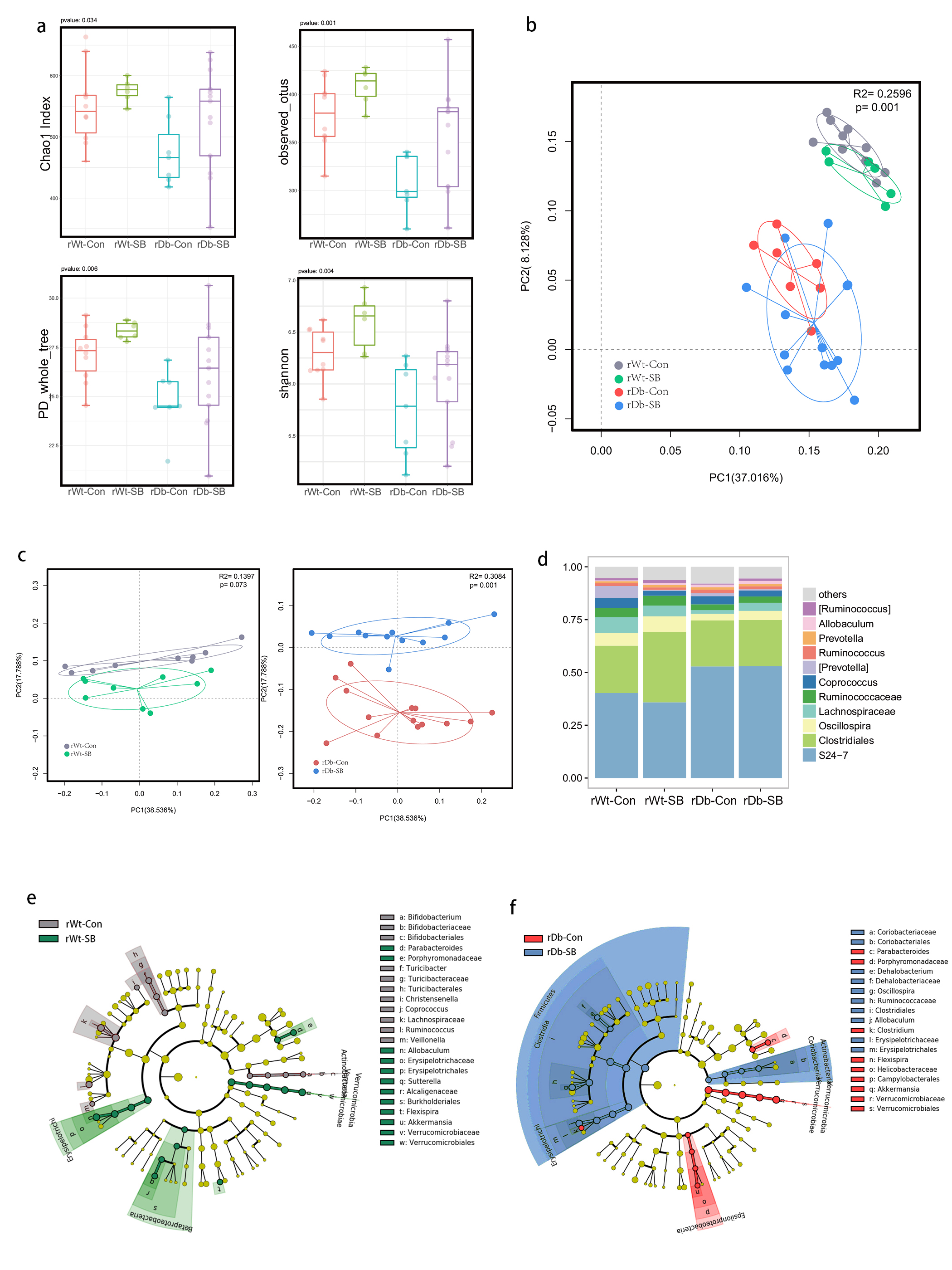
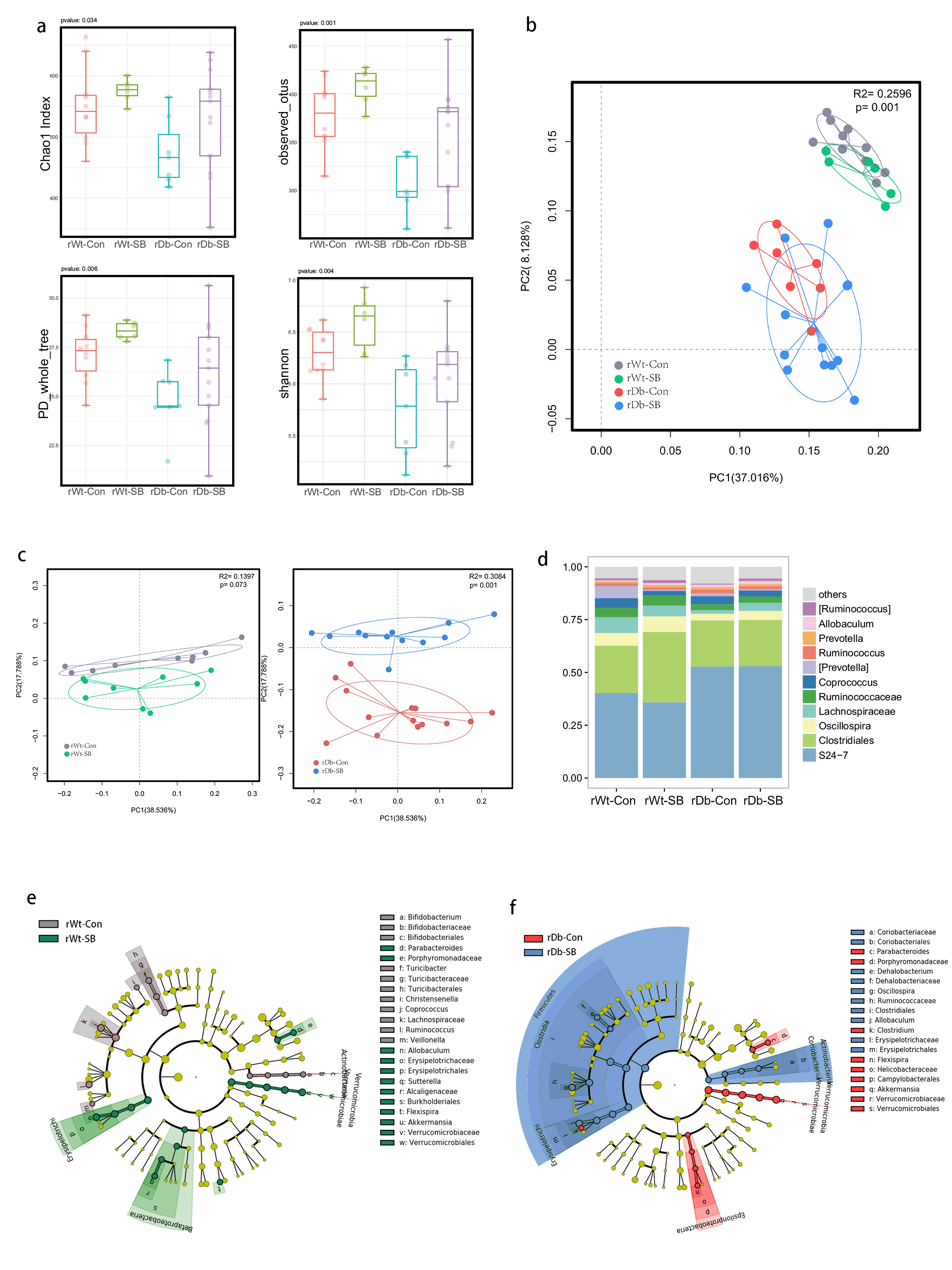
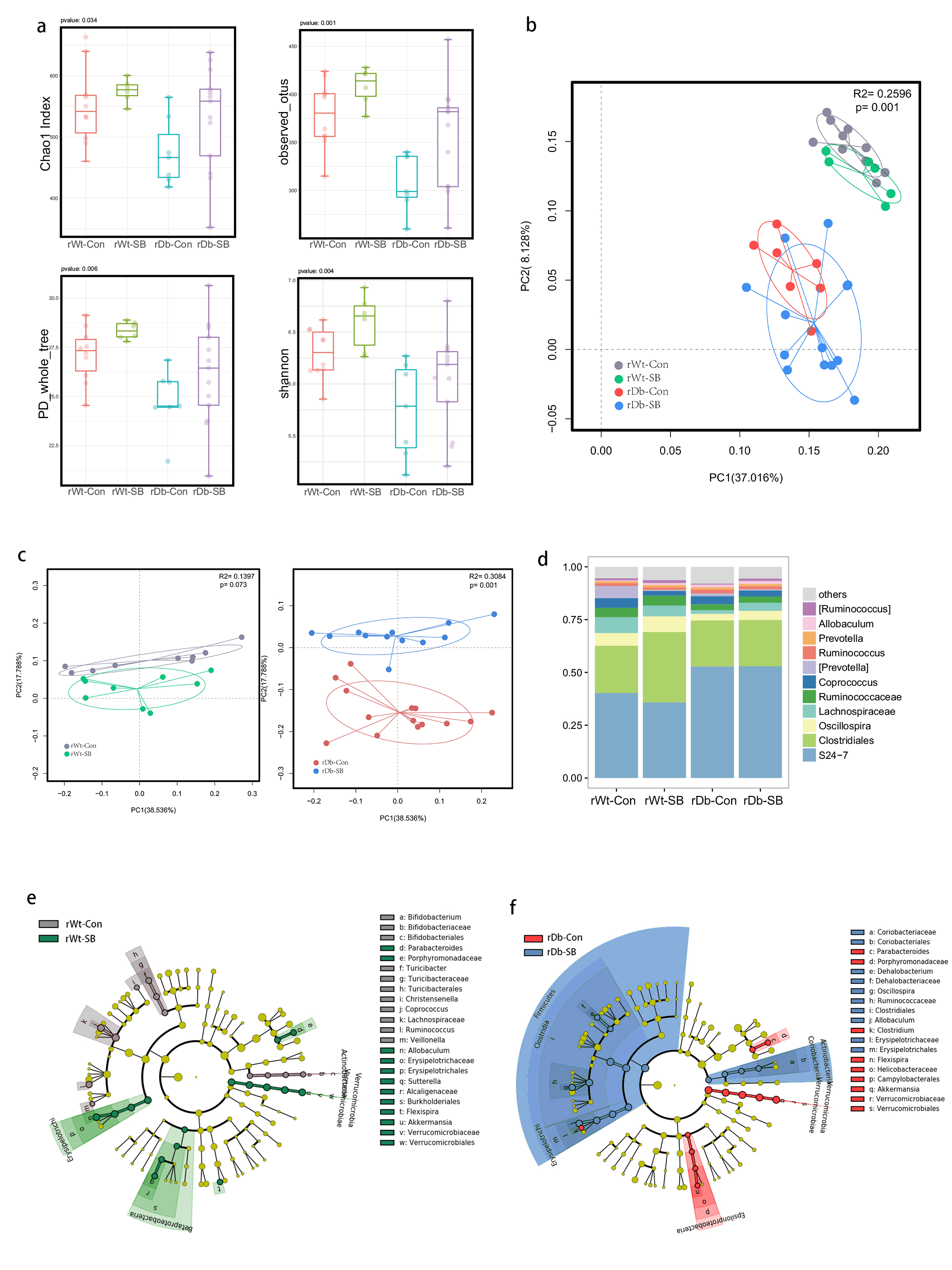
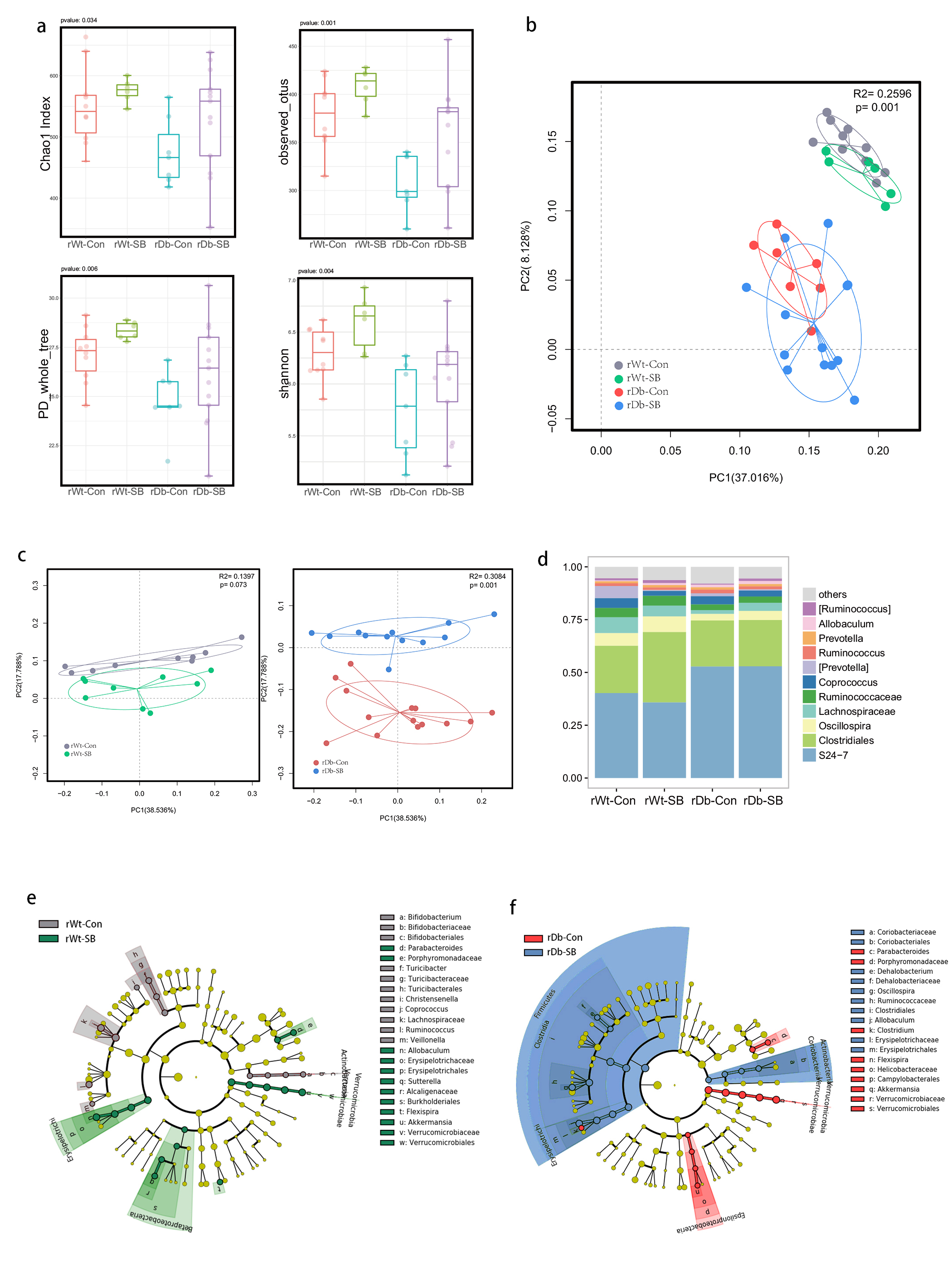
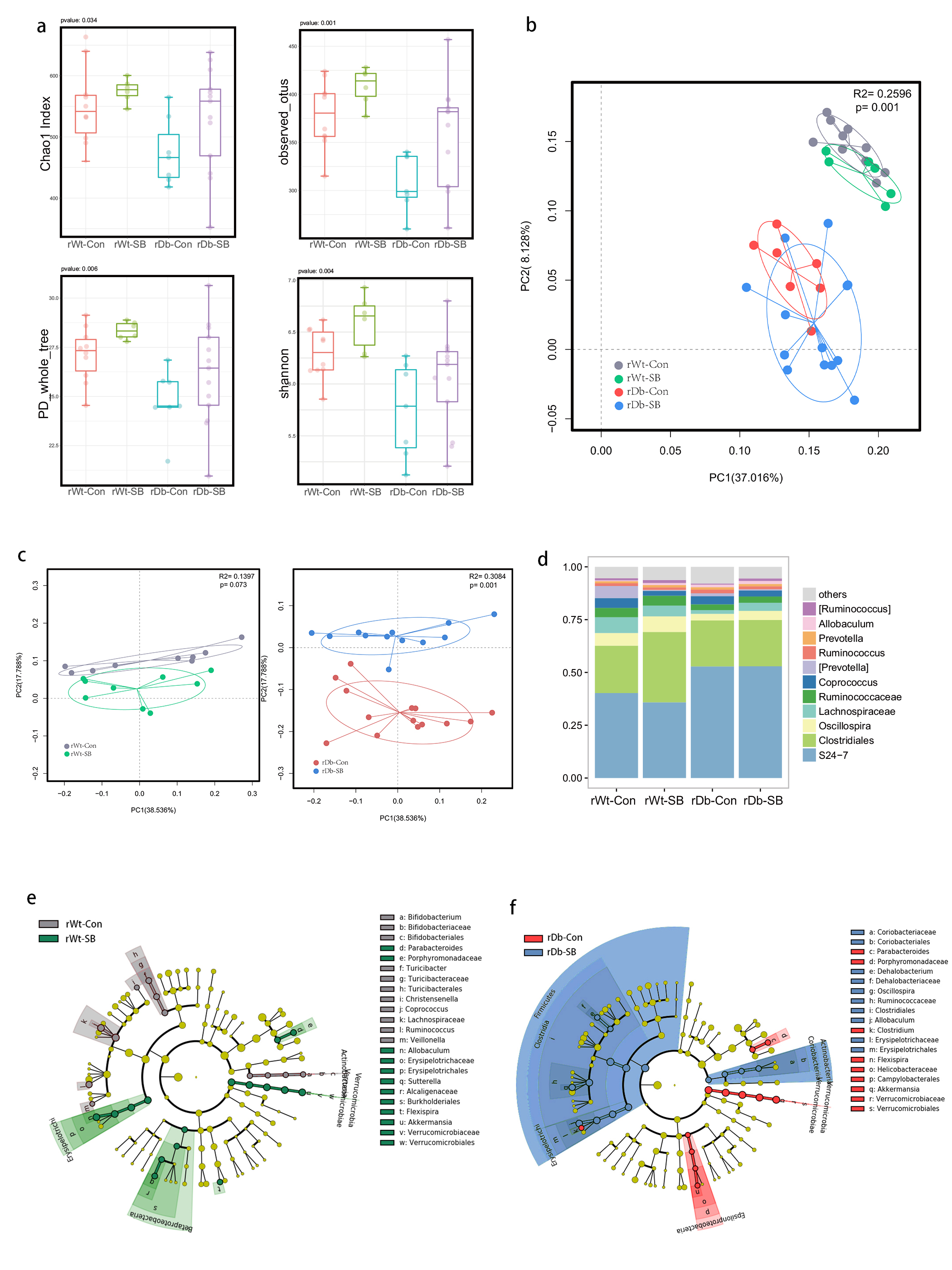
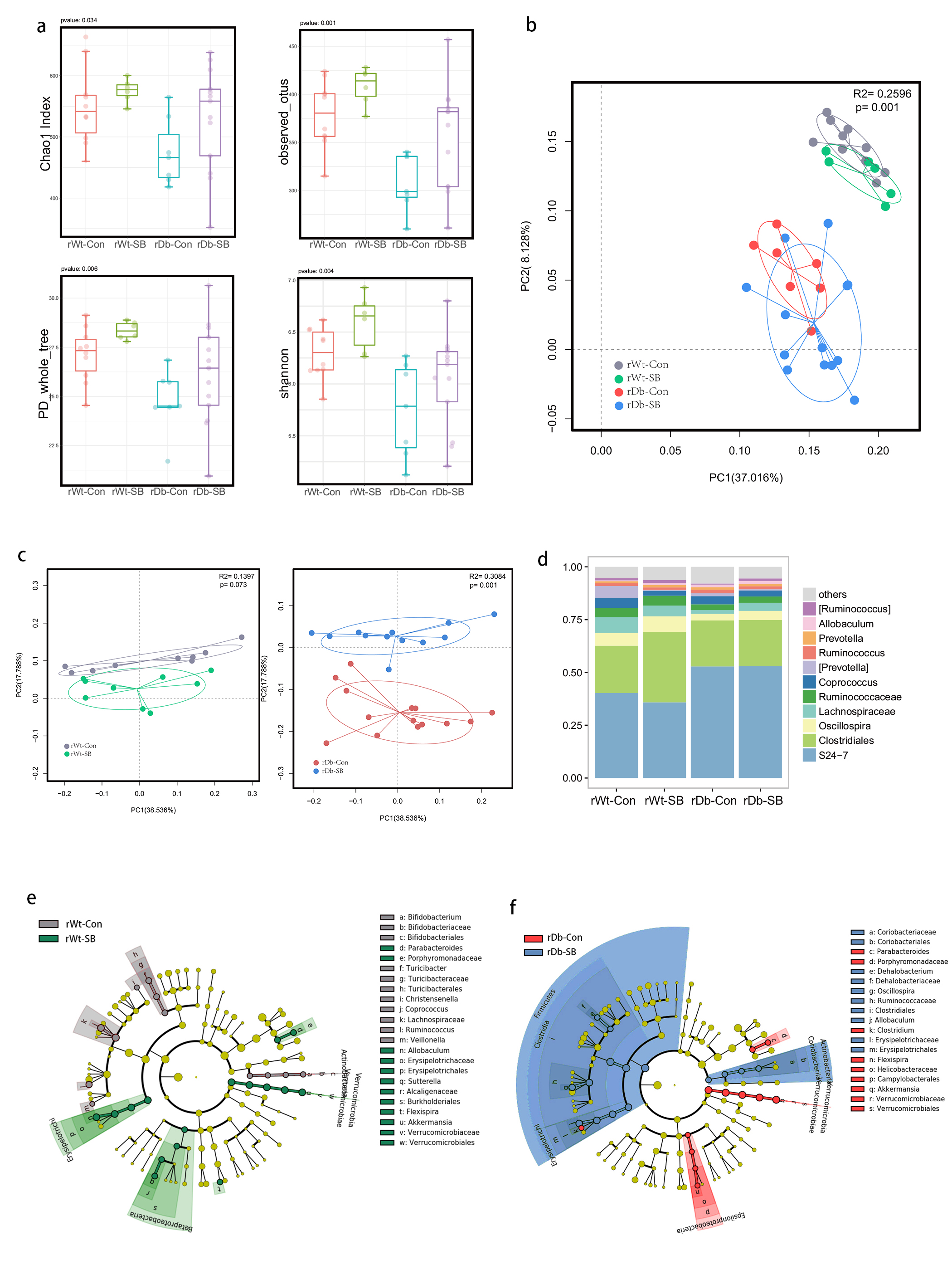
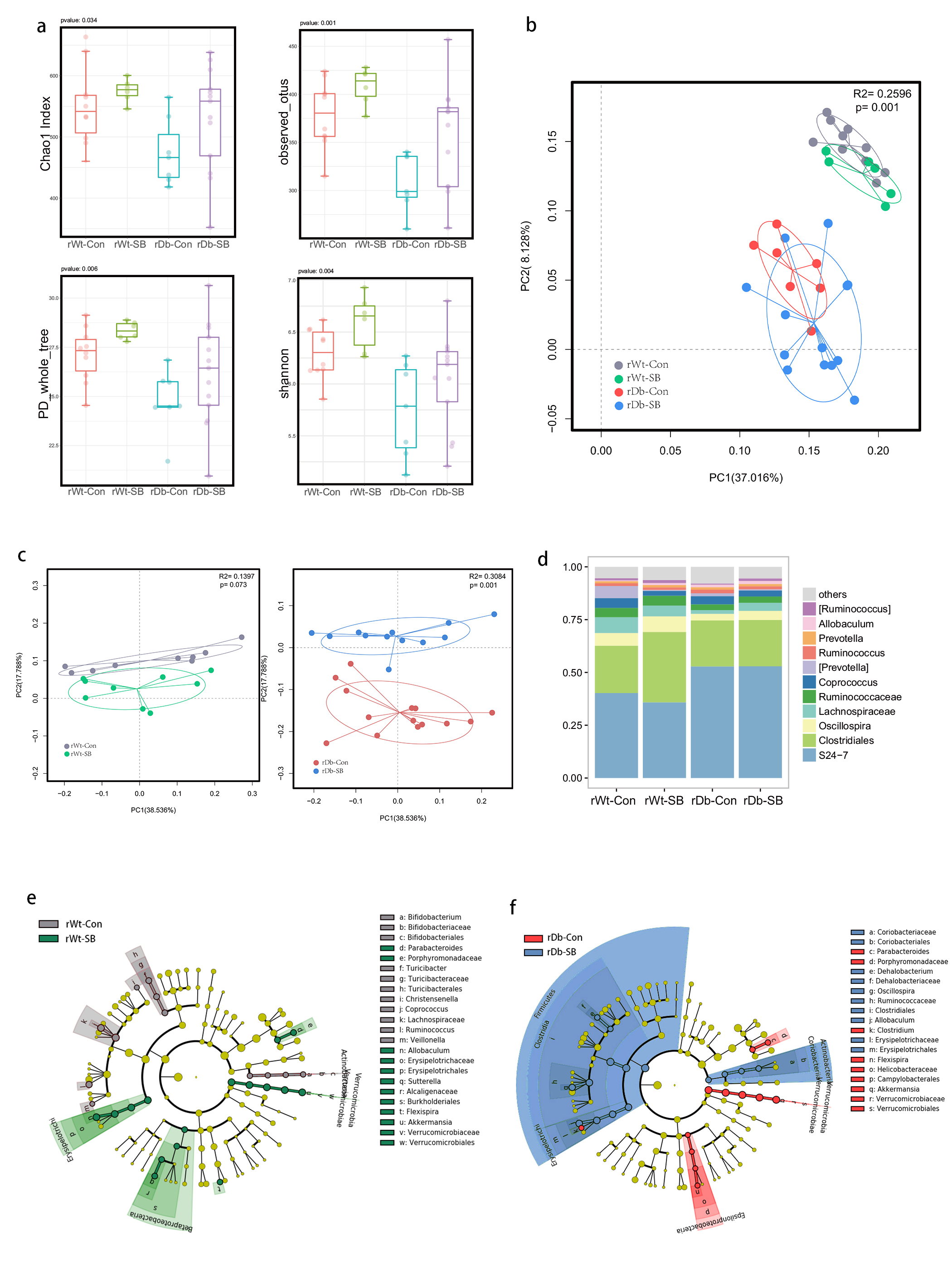
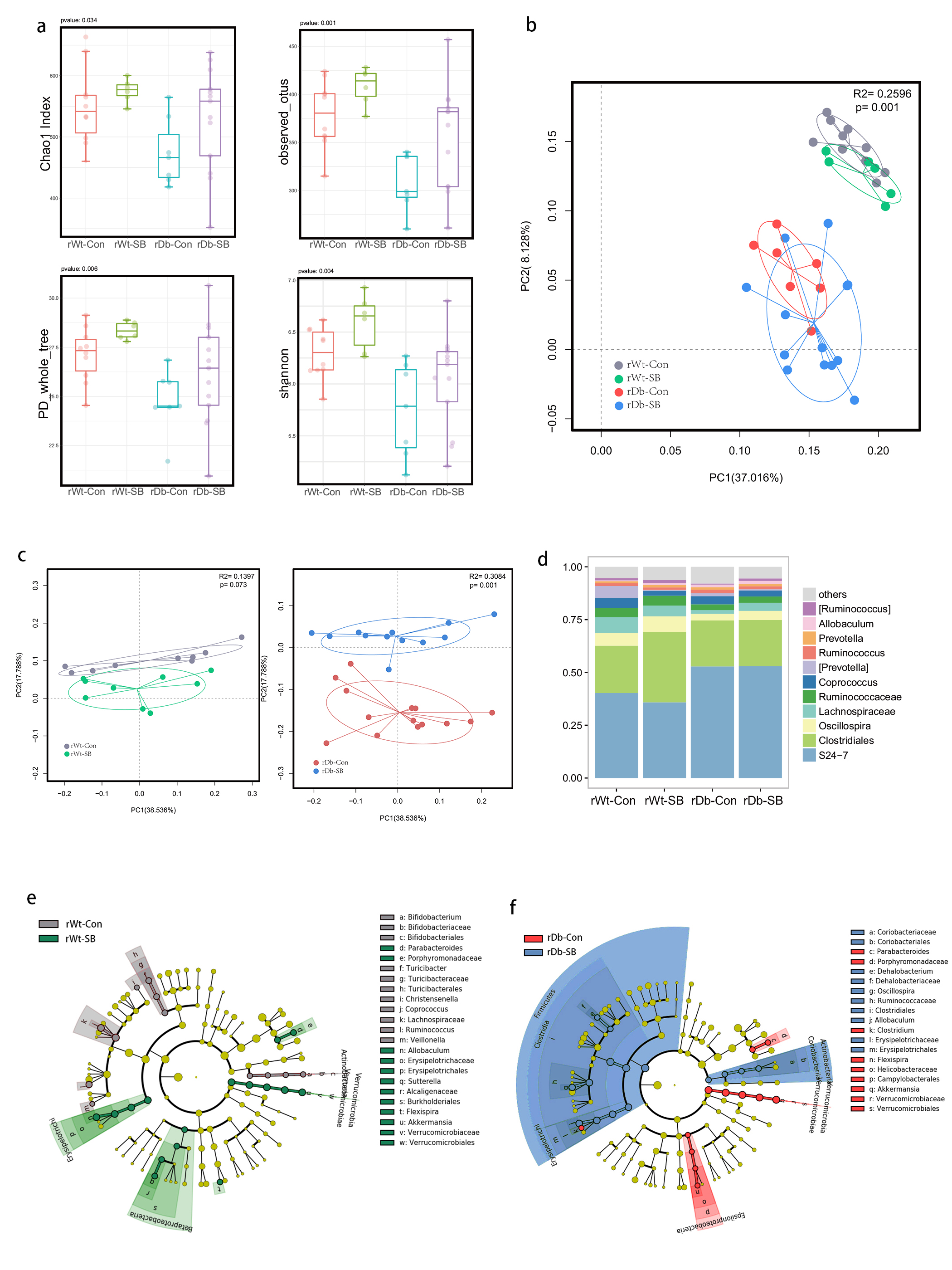
In this study, we found that the gut microbiota profile of Db mice was distinct from that of Wt mice. Several studies have shown that the gut microbiota might be an important contributor to the development of T2D. Ridaura et al. discovered that the obesity-associated metabolic phenotype can be transmitted from humans to mice by FMT [28]. In males with metabolic syndrome, FMT from lean male donors results in a significant improvement in insulin sensitivity, increased gut microbiota diversity and a remarkable increase in butyrate-producing bacteria [29]. However, in this study, FMT from Db mice had no effect on diabetes-related parameters, possibly due to the short period of transplantation. Studies by both Qin et al. [8] and Karlsson et al. [30] showed that T2D subjects exhibit reduced abundances of Clostridiales bacteria, such as Roseburia and Faecalibacterium prausnitzii, which are known as butyrate-producing bacteria. Thus, supplementation with butyrate might elicit favorable effects on diabetes-related parameters. In this study, we found that SB supplementation reduced food/water intake, fasting blood glucose levels and serum TG levels in Db mice. These data are consistent with those of Mollica et al., who found that oral administration of butyrate improves insulin sensitivity, and reduces serum TG levels and energy expenditure in diet-induced obese mice by enhancing mitochondrial function [31].
We also found that SB supplementation shaped the gut microbiota of Db mice, with SB-treated mice exhibiting a relatively higher abundance of butyrate-producing bacteria than NaCl-treated mice. Notably, in line with data published by Zhou et al. [19], we observed that SB treatment significantly increased the abundance of Christensenellaceae, a recently described family member of the phylum Firmicutes. Christensenellaceae have been shown to be enriched in individuals with a normal body mass index (BMI) compared to obese individuals with a BMI over 30 [32]. Moreover, increases in Christensenellaceae have been demonstrated following diet-induced weight loss [33]. A recent study found that a higher abundance of Christensenellaceae was positively associated with a lower cardiometabolic risk score in a cohort of 441 Colombians [34]. Moreover, Christensenellaceae has also been found to be associated with healthy glucose metabolism [35, 36]. However, the mechanism underlying its negative association with metabolic symptoms remains to be elucidated. Future studies focusing on Christensenellaceae might help gain insight for the treatment of T2D.
T2D is a well-known comorbidity that aggravates brain injury after ischemic stroke. The pathophysiological mechanisms by which T2D exacerbates brain injury after stroke have not been fully elucidated. In recent years, tremendous progress has been made in identifying the bidirectional interactions that occur between the gut and the brain, namely, the gut-brain axis. A series of provocative studies have suggested a prominent role for the gut microbiota in ischemic stroke [22–24]. Thus, we aimed to seek answers from the gut. A previous study revealed that Clostridium butyrium attenuates ischemic stroke injury in diabetic mice via modulation of the gut microbiota [37]. However, this study does not discount the effect of the blood glucose level on ischemic stroke injury, as treatment with Clostridium butyrium significantly alleviates hyperglycemia, which is a strong risk factor for poor stroke outcome. Admission hyperglycemia is associated with poorer stroke outcome possibly because it exacerbates ischemic injury by disturbing recanalization and worsening reperfusion injury [2]. Studies also suggest that inflammation may contribute to ischemic stroke concomitant with T2D [38]. Thus, we used FMT and MCAO models of stroke to address the causative relationship between the gut microbiota and stroke injury independent of the regulation of glycemia. As a result, mice that underwent FMT from SB-treated Db mice exhibited lower neurological deficit scores and a smaller cerebral infarct volume than mice that underwent FMT from NaCl-treated Db mice. Importantly, we found that the rDb-SB group had a higher abundance of butyrate-producing bacteria and a higher level of butyrate, which appear to be at least partially responsible for protection against ischemic brain injury, than the rDb-Con group. Benakis et al. found that intestinal IL-17-producing γδ T cells travel from the gut to the brain meninges following ischemic stroke to influence stroke injury [22]. Gut microbes have been demonstrated to induce CD4+ T cells to differentiate into proinflammatory T cells or anti-inflammatory regulatory T cells [39–41]. Notably, SCFAs regulate leukocyte trafficking from the gut to other tissues, such as the uvea [42]. Moreover, SCFAs have been reported to regulate microglial homeostasis [43] and are important for the development and maintenance of the BBB [44]. In the current study, we observed that the rDb-SB group exhibited less microglial activation, neuronal loss and apoptosis in the ipsilateral cortex and hippocampus region of the ischemic hemisphere than the rDb-Con group. Our findings indicate that the gut microbiota in T2D per se has a great impact on ischemic stroke injury and can be influenced by SB treatment.
Ischemic stroke is often accompanied by BBB disruption. BBB dysfunction triggers vascular edema and creates an inflammatory environment, ultimately resulting in neuronal death and brain damage [45]. Therefore, maintenance of BBB integrity is thought to be key for protecting the brain from ischemia/reperfusion injury. A major finding of the present study was that modulation of the diabetic gut microbiota by SB attenuates cerebral ischemia by protecting the BBB. Some of the most important proteins that affect BBB permeability are tight junction proteins. Our data revealed that degradation of Occludin and Claudin-4 was relieved in the rDb-SB group compared with the rDb-Con group. Moreover, we observed that the rDb-Con group experienced the severest endothelial glycocalyx degradation and the highest serum concentrations of its components, i.e., the HS and syndecan-1. Interestingly, the endothelial glycocalyx in the rDb-SB group was mildly damaged. The endothelial glycocalyx, a gel-like layer that covers the vascular endothelial surface and floats into the lumen of the vessels, plays a critical role in vascular integrity and cardiovascular homeostasis [46]. In our previous report, we discovered that endothelial glycocalyx degradation leads to increased BBB permeability in a rat model of asphyxia cardiac arrest [47]. Collectively, these data indicate that SB modulates the gut microbiota of T2D mice and that this modulatory effect affords neuroprotection against cerebral ischemia/reperfusion injury. However, future studies are necessary to determine the underlying mechanisms.
We acknowledge that this study has several limitations. First, although the Db mouse model used in the present study is the most widely used model of T2D, the monogenetic underpinnings of this model are not a perfect representation of T2D in humans. Different T2D mouse models, e.g., db/db mice and streptozotocin-induced diabetic mice, may exhibit different gut microbiome profile. Second, for FMT, we used the entire fecal content, which includes bacteria, fungi, viruses, metabolites from bacteria and undigested food. Therefore, it is not clear which component of the feces exerts effects against ischemic stroke injury. The elevated concentration of butyrate in the SB-treated group may have partially resulted from SB in the drinking water. Third, even though we confirmed that SB remodels the gut microbiota of Db mice to afford neuroprotection against cerebral ischemia injury, we did not explore the underlying mechanism. It is believed that intestine-derived T cells play a vital role in brain injury after stroke, and future studies investigating the immune status of T2D treated with SB might shed light on the underlying mechanism.
In conclusion, our findings demonstrate that SB is sufficient to shape the gut microbiota of Db mice and elevate the fecal concentration of butyrate, which alleviates diabetes-related symptoms. Restoration of the gut microbiota of T2D mice by SB can protect the BBB and reduce cerebral infarct volume. This study provides the first experimental evidence that the microbiota composition can be therapeutically exploited to protect against cerebral ischemia injury in patients with T2D. However, future studies are required to elucidate the underlying mechanism.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}