Microorganisms that thrive in hostile environments demonstrate a variety of adaptive strategies, most notably biofilm formation (Fröls, 2013; Yin et al., 2019). A multitude of bacteria and fungi can shift between planktonic and multicellular biofilm communities, of which approximately 80% exist in the environment as biofilms (Donlan, 2002; Ramage et al., 2012). Fungal biofilms can significantly amplify the resistance to environmental factors (Manganyi et al., 2015). Therefore, studying biofilm formation is essential for understanding the behavior and structural changes of plant pathogenic fungi thriving under hostile conditions.
Several microbes have evolved mechanisms of forming and encapsulating themselves, either singly or in communities, within outer coating structures known as biofilms. These biofilms facilitate microbes to stick to one another or to surfaces (Motaung et al., 2020; Penesyan et al., 2021). The material that encases cells and binds them together is known as the extracellular matrix (ECM) (López et al., 2010), a hallmark of biofilm formation by fungal pathogens (Costa-Orlandi et al., 2017; Liu et al., 2022; Mitchell et al., 2016a, 2016b; Zarnowski et al., 2021). This matrix is composed mainly of a polysaccharide biopolymer along with other components such as proteins, nucleic acids, lipids, and exopolysaccharides (Branda et al., 2005; Motaung et al., 2020). In particular, exopolysaccharides can vary significantly based on growth requirements, nutrient composition, (López et al., 2010), temperature, and pH. Biofilm formation offers cells the ability to withstand difficult conditions and the possibility of stretching out and finding new locations to inhabit (Hall-Stoodley et al., 2004).
Formation of biofilms is implicated in having negative consequences to healthcare, biotechnological, and agricultural industries. This is partly due to the limited understanding of the mechanisms underlying biofilm adaptation to different microenvironments (Cámara et al., 2022; Mishra et al., 2020). As biofilms form, they can interact with surfaces and result in the corrosion, deposition, and microbiological degradation of materials. This can be a challenge to the healthcare sector, as biofilms can be difficult to remove from medical equipment such as dentures, endotracheal tubes, orthopedic prostheses, cardiac pacemakers, catheters, and heart valves. Their ability to persist is due to antifungal resistance of the three-dimensional (3D) biofilm structure which in turn escalates biofilm-related clinical infections, thereby increase the cost of treatment (Bryers, 2008; Coad et al., 2016; Desai et al., 2014; Donlan, 2001; Wi & Patel, 2018). The negative outcomes of biofilm formation in biotechnological environments can include biofouling, corrosion, and a decrease in the quality of drinking water (Mishra et al., 2020). Similarly, the same challenges may occur in agriculture when the necessary machinery and tools are contaminated with harmful fungal biofilms, making it a challenge to remove them. Biofilm colonization on farming equipment has the potential to contaminate unaffected fields with unique biofilm-generated propagules, some of which may have acquired resistance to fungicides (Motaung et al., 2020; Peremore, 2022b; Shay et al., 2022). In agro-based industries namely aquatic, meat, dairy, and food processing plants, biofilm formation contaminates food and food sources, which poses a threat to food safety (Toushik et al., 2022).
The environment including pH levels, temperature, nutrient availability, and hydrodynamics impact on biofilm formation. These conditions influence the composition and structural development of biofilms by affecting the enzymes and regulatory processes involved in biofilm maturation and dispersal (Flores-Vargas et al., 2021; Goller & Romeo, 2008). For instance, changes in nutrient composition can result in alterations within ECM produced by microbial communities and this affects the stability and protective functions of biofilms (Goller & Romeo, 2008; Mirghani et al., 2022). Furthermore, variables such as the presence of specific ions and water flow velocity can influence the 3D structure of biofilms, impacting their resilience and interaction with the surrounding environments (Balcázar et al., 2015; Flores-Vargas et al., 2021). Understanding these environmental factors is critical for understanding how biofilms grow and adapt under various conditions.
Although research on phytopathogenic biofilm formation and its effects on the microbial environment in the agricultural sector exists (Peiqian et al., 2014; Peremore, 2022b; Ratsoma et al., 2023; Shay et al., 2022; Tyzack et al., 2023), but it is not as extensive as with other sectors. Very recently, formation of biofilm within plant fungal pathogens namely; Fusarium verticilloides, Fusarium circinatum, Fusarium oxysporum f. sp. cucumerinum, Fusarium graminearum and Zymoseptoria tritici has been confirmed (Motaung et al., 2020; Peremore, 2022b; Ratsoma et al., 2024; Shay et al., 2022). From these studies, there is consensus that biofilms provide protection from host defences and extreme environments.
Exserohilum turcicum, the causal fungal pathogen of northern leaf blight (NLB), the most prevalent global foliar disease of maize in sub-Saharan Africa (Nsibo, 2019). Exserohilum turcicum survives on infected maize residues and spreads either by wind or splash under favourable weather. (Wise, 2011; Yang et al., 2017). The pathogen is implicated in causing up to 75% yield losses if not managed effectively and hence is a major threat to Africa’s food security (Berger et al., 2020). Control measures currently used to manage infections include crop rotation, deep tillage, NLB-resistant maize cultivars, and fungicides (Human et al., 2016; Kotze, 2020). Exserohilum turcicum is known to exhibit high genetic diversity in South Africa (Human et al., 2016; Nieuwoudt et al., 2018) being driven by several evolutionary forces namely; migration, sexual reproduction, and/or mutation (Haasbroek, 2014). The high genetic diversity found within the E. turcicum populations may lead to the emergence of strains that are more resistant to fungicides and other management strategies in the long run. Additionally, sexual recombination among strains can facilitate the exchange of genetic material, thus giving rise to more virulent strains in nature and adding complexity to the strategies for managing fungicide resistance (Bankole et al., 2023; Human et al., 2016; Nieuwoudt et al., 2018).
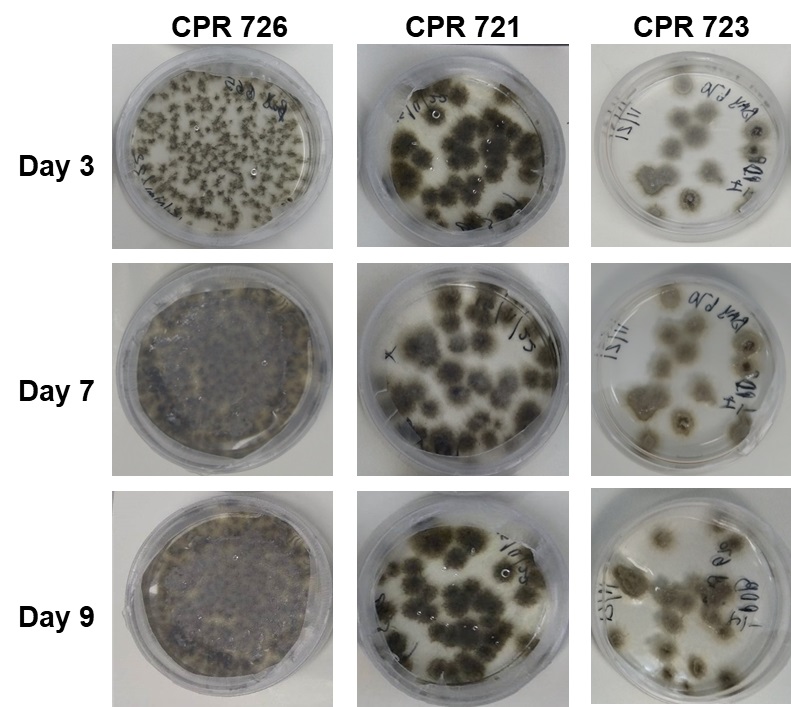
To our knowledge, no study has characterized biofilm formation in E. turcicum within a single strain or at a population level. The lack of this knowledge derails the development of more effective control strategies of the pathogen as well as the effort to understand how it behaves in nature. Therefore, the aim of this study was to screen and characterize biofilm formation in E. turcicum strains and determine the role of ECM production in response to abiotic stresses. The study explored several scientific techniques (plate and microscopic analysis (SEM, CLSM and TEM)) to screen and characterize E. turcicum biofilm formation and its environmental response at the morphological and metabolic levels.
{kind=link}