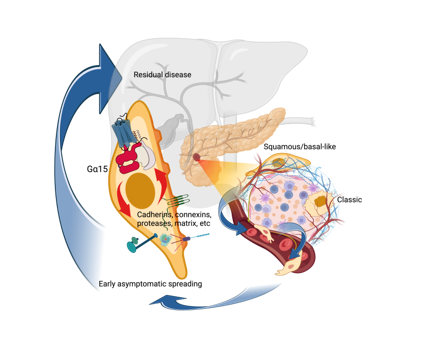
3.1 GNA15 expression in cancer cells is associated with squamous features
Squamous cells are absent in normal pancreas but emerge when aberrant ductal-to-squamous transition alters cell fate and drives oncogenesis 19. GNA15 is ectopically induced in pancreatic neoplasia and is associated with more aggressive oncogenesis 10. A survey of human gene expression data for the multiple cancers of TCGA-ICGC shows that GNA15 expression is strictly associated to the five squamous cell carcinomas (Fig. 1A).
Within this subset of malignancies, GNA15 expression positively correlates with the expression of TP63 23 (Fig. 1B) a master regulator of the normal squamous lineage and the transformed basal-like/squamous subtype of PDAC 24. Consistently, in TCGA-PAAD cohort, TP63 and GNA15 are overexpressed in the basal-like/squamous transcriptional subtype (Fig. 1C). The basal-like/squamous subtype has also been defined as “glycolytic”, in contrast to the “lipogenic” ‘classical/progenitor’ type 3,25. We find GNA15 expression is proportional to hypoxia scores (Winter’s 20, Ragnum 21 and Buffa 22) based on markers like GAPDH (Fig. 1D) and others specific of the glycolytic component of the basal-like/squamous subtype (Supplementary Table 1). The hypoxia score was reported to be similar in parent lesions in the primary tumor versus matched daughter metastatic lesions and it correlated with a higher frequency in bi-allelic loss of the oncosuppressor TP53 independently of the microenvironment or a particular clinical or genetic feature 26. Recently the loss of TP53 heterozygosity was hypothesized to characterize pre-tumor cells predisposed to progress towards subclonal evolution 27. Thus, hypoxia-adaptation is likely to represent an inherent characteristic of the basal-like/squamous subtype, rather than a transient response to external conditions. Epigenetically driven GNA15 expression could be part of this early adaptation; it is not correlated with the presence of most prominent driver mutations in PDAC (Supplementary Fig. 1) but is directly correlated with loss of heterozygosity of TP53 and inversely correlated to the expression of the oncosuppressor SMAD4 (Fig. 1E, Supplementary Table 2) reported to be significantly reduced in the basal-like/squamous subtype 4.
To better understand the significance of GNA15 ectopic expression we analyzed TCGA-PDAC for co-expressed genes.
3.2 GNA15 expression is associated with an expression signature related to cell-cell interactions
Most genes co-expressed with GNA15 (Fig. 2A) have been previously reported to play a role in PDAC development (Supplementary Table 2). These associations were validated by RNAseq (Fig. 2B) and by RT-PCR (Fig. 2C) in 22 PDAC patient-derived xenografts (PDX).
Strengthening GNA15 expression as an indicator of poor prognosis 10, all positively correlated genes predicted poorer prognosis, opposite to negatively correlated genes (Supplementary Fig. 2). Gepia 2.0 analysis of neoplasia datasets present in TCGA reveals that GNA15 signature predicts earlier relapse in PDAC as compared to other cancers. The four genes most highly correlated to GNA15 expression encode transmembrane proteins. These genes comprise a PDAC specific restricted signature with a significant hazard ratio (Fig. 2D) that predict disease free survival comparable to a larger signature of 200 co-expressed genes with a Pearson score over 0.5, Fig. 2E and Supplementary Fig. 2B. These four genes could thus serve as prognostic biomarkers.
The gene most highly correlated with GNA15 expression is tumor-associated calcium signal transducer 2 (TACSTD2), the homolog of EpCAM that encodes the trophoblastic cell surface antigen 2 (Trop-2). Histological analysis confirmed the co-localization in neoplastic lesions (Supplementary Fig. 3A) although GNA15 is more restricted to neoplastic cells. TACSTD2 is also expressed by epithelial cells 28, particularly squamous, and overexpressed in several adenocarcinoma, particularly squamous cancers (Supplementary Fig. 3B). TACSTD2 over-expression correlates with poor prognosis in pancreatic and other carcinomas, including up to 95% of triple negative breast cancer where Trop-2 represents the target of the FDA-approved antibody-drug conjugate Sacituzumab govitecan (IMMU-132) 28.
We previously evaluated the functional impact of reducing Gα15 expression in the PT45 PDAC cell line. GNA15 was downmodulated by expressing shRNA or completely knocked out by CRISPR/Cas9 10. Using the same models, here we analyzed the influence of Gα15 on the abundance of Trop-2 by flow cytometry and western blot. In all cases, Trop-2 expression increased when GNA15 was depleted (Fig. 3A, Supplementary Fig. 4A).
Trop-2 and Gα15 are involved in Ca2+ and PKC signaling, carcinoma cells motility 10,28, PDAC progression and invasiveness 10,29,30. Several other genes in the signature encode for membrane proteins implicated in PDAC cell motility and reset contacts with the microenvironment (Supplementary Table 2). The GJB3, CDH3, CDCP1 genes encode connexin 31, p-cadherin and CUB Domain Containing Protein 1 (CDCP1) respectively. The latter prevents adhesion through integrin signaling 31, is phosphorylated by Src in metastatic cancers and its interaction with PKC contributes to anoikis resistance 32. CDCP1 was identified by a combination of parallel proteomics and CRISPR screenings aimed to identify KRAS induced cell surface proteins 33.
To explore if GNA15 signature proteins regulate homotypic cell-cell interactions we compared cell aggregation in PT45 clones with or without GNA15. Loss of GNA15 expression increased the ability of cells to form aggregates (Fig. 3B) suggesting Gα15 may promote dispersal of transformed cells.
3.4 GNA15 knockout affects expression of genes instrumental to cell spreading
We next analyzed the impact of GNA15 knock-out (Supplementary Fig. 4B) on the PANC-1 cells transcriptome (Fig. 4A). Gene ontology of RNAseq results showed that cell adhesion and migration were the most significantly impacted by GNA15 depletion (Fig. 4B).
Plasminogen Activator, Urokinase Receptor gene (PLAUR) emerged among the most downmodulated genes, consistent with its role in promoting cell invasion 34. Interestingly, analyzing TCGA-ICGC data for gene expression in neoplasia in various tissues, shows PLAUR is most highly correlated with GNA15, directly followed by the gene encoding its ligand, PLAU. A key role for PLAUR in regulating the shift between single cell tumor dormancy and proliferation has been attributed to PLAUR induction by integrins, resulting in the propagation of signals from fibronectin (FN1) through the EGF-receptor, ERK, and p38 signaling (see Discussion). FN1 was also one of the genes most significantly downregulated by GNA15 knockout in PANC-1 cells. PLAUR and FN1 gene expression levels were validated by TaqMan PCR (Fig. 4c). Additional genes affected by GNA15 knock out support this hypothesis. Of particular interest are the downmodulation of pro-migratory insulin-like growth factor binding proteins (IGFBPs) 1 and 6 35 and the upregulation of annexin A6 (ANXA6), which prevents migration and invasiveness of squamous epithelial cells 36.
3.5 Gna15 facilitates the development of PDAC precursor lesions in KC mice
Studies in PDAC cell lines showed that KRAS affects the function of membrane proteins to reduce adhesion and promote invasiveness 33. KRAS mutation is an early event in the etiology of several cancers in humans. Oncogenic KRAS alleles are found in precancerous PanIN-1A (25%) and PanIN-1B (38%) lesions, and predominate in PDAC (85%) 37. Since GNA15 is ectopically expressed by the same lesions 10, we sought to test if Gα15 contributes to the initiation of pancreatic neoplasia leading to PDAC. A mouse model that expresses oncogenic LSL_KrasG12D in all cells of the pancreas under control of Pdx1::Cre (KC) from embryonic day 9 (e9) was crossed with a whole body null allele of Gna15 38 to obtain KC;Gna15-/- mice (KCG).
Pancreas development and adult function are normal in Gna15 KO mice 38. Analysis was performed on KCG and KC pancreata obtained from either sex at about 40 days of age. At this early stage of initiation, the lesions in both lines of mice are distinct, relatively small, non convoluted and spheroidal 39. Evaluation of serial sections of pancreas from at least 3 different levels separated by 150 µm showed that individual early preneoplastic lesions were morphologically similar but fewer in KCG compared to control KC pancreata (Supplementary Fig. 5 and Supplementary Table 3, p < 0.005). Detailed analysis of a representative H&E section obtained for each mouse quantified all lesions in non-overlapping fields. In total, KC pancreata had a 3-fold greater lesion burden than KCG, displayed higher proportions of fields with single or multiple lesions, and correspondingly fewer negative fields than KCG (61% vs. 86%, Fig. 5).
The majority of KCG lesions occurred individually in well separated fields although two pairs of neighboring fields had two or more lesions (KCG#10 and #12, Fig. 5). By contrast, most lesions in KC pancreata occurred in arrays of 2–6 contiguous fields, and these contained almost all of the fields with more than one lesion. KC had 2-fold higher ratio of fields with one lesion, 8-fold more with two lesions, 10-fold or more with three or four lesions compared to KCG (Supplementary Table 3). Note that nearly every multi-lesion field is neighbored by an empty field in both KC and KCG pancreata, suggesting local signaling drives metaplasia evoked by oncogenic Kras. Because oncogenic Kras is the only driver of neoplasia in the KC model, the results are consistent with the hypothesis that Gα15 ectopic signaling precedes additional driver mutations. Our mouse and human data indicate that regardless of mutations that neutralize oncosuppressors typically driving PDAC, Gα15 signaling intensifies oncogenic KRAS-dependent initiation of preneoplastic lesions in pancreas.
{kind=link}