Ket is one of the anesthetic drugs that have different effects on mood and behavior in different doses. In low doses, it has antidepressant effects, and in higher doses, it causes excessive movement and behavior, such as becoming delirious. As seen in this study Ket administration in higher doses can be a promising animal model for mania. Ket induced hyperactivity in the open field test. Moreover, Ket administration increased the activities of the TCA cycle enzyme (MDH and SDH) and mitochondrial respiratory chain complex (I, II, III, and IV). Li was able to reverse almost totally these effects.
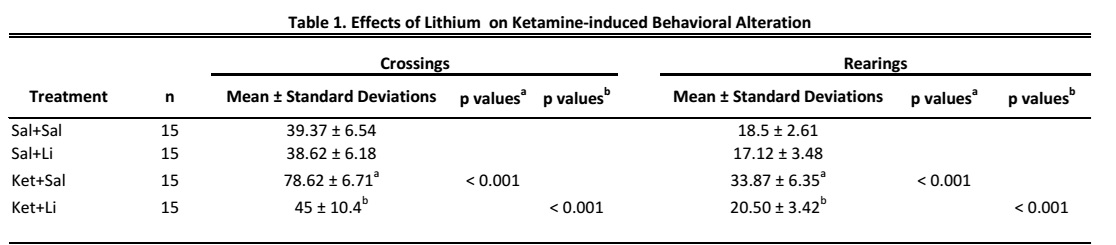
Ketamine, a dissociative anesthetic, in rodents induces psychotic behavior as hyperlocomotion (Canever et al., 2010; De Oliveira et al., 2011). Moreover, Ricker et al. (2011) showed that Ket therapy was able to induce prolonged mania associated with psychotic symptoms in a reflex sympathetic dystrophy patient. Therefore, our behavioral results corroborate with recent preclinical and clinical studies, indicating that Ket can induce an animal model of mania (Ghedim et al., 2012; Chaves et al., 2020). Ghedim et al. (2012) demonstrated that Li can reverse and prevent Ket-induced hyperlocomotion by causing changes in the membrane's excitability, which may be related to its antimanic effect. Therapeutic concentrations of this drug practically do not present psychotropic effects in normal individuals, not behaving as a sedative, depressant, or euphoric, a characteristic that distinguishes it from other psychotropics (Ghedim et al., 2012; Chaves et al., 2020).
We also evaluated the enzymatic activity of the TCA cycle (MDH and SDH), and the results demonstrated that Ket induced an increase in the activity of these enzymes. Ket is involved in behavioral and biochemical changes, such as increased mobility, oxidative stress associated with mitochondrial dysfunction, and changes in brain-derived neurotrophic factor levels observed in BD patients. Feier et al. (2011) showed that the administration of m-amphetamine, an animal model of mania, causes increased Citrate Synthase (CS) and SDH in the prefrontal cortex, amygdala, hippocampus, and striatum, and Li and valproate were able to reverse this effect. Ket induces an animal model of schizophrenia (Canever et al., 2010; De Oliveira et al., 2011). In addition, a study has shown in the post-mortem brain of schizophrenic patients the activity of increased SDH and MDH (Bubber et al., 2011). Although Li reverses these effects by altering the transport of cations across cell membranes and in nerve and muscle cells. Furthermore, due to its similarity with other elements (such as sodium and potassium), lithium can influence the reuptake of serotonin and/or norepinephrine and allows it to inhibit the second messenger systems involved in the phosphatidylinositol cycle (Freitas et al., 2010; Valvassori et al., 2014).
The animal models of disorders in humans must meet three sets of criteria, such as construct, face and predictive validity BD is accompanied by recurrent manic and depressive episodes (Goodwin and Jamison, 2003). Manic episodes are characterized by irritable mood, psychomotor activation, reduced sleep requirements, and excessive involvement in potentially problematic behavior (El-Mallakh et al., 2003). One of the few symptoms of this disorder that can be reproduced in animal models is hyperactivity, which was observed in this study. In addition, Li was used to test the efficacy of the anti-manic treatment in this animal model, and we observed that Li completely reversed the hyper-compliance induced by Ket. Therefore, the results found in this study showed that the animal model of mania presented in this study may induce behavioral changes and response to Li treatment, showing similarities to BD, indicating that this animal model induced by Ket presents face and face validity predictive.
In this work, the activity of the complexes I, II, II-III, and IV of the mitochondrial respiratory chain in this animal model was also evaluated. The results indicate that Ket increased the activity of all mitochondrial complexes. Pre-clinical studies show that the activity of these complexes is decreased in animal models of mania induced by m-AMPH and d-AMPH in the hippocampus, striatum, and prefrontal cortex, and Li was able to reverse these effects (Valvassori et al., 2010; Feier et al., 2013). Some preclinical and clinical studies agree with the results of this study. Freitas et al. (2011) showed that intracerebroventricular administration of ouabain, an animal model of mania, induces the increased activity of mitochondrial complexes I, II, II-III, and IV in the prefrontal cortex of rats. A clinical study also corroborates these data, as it shows increased mRNA levels of the mitochondrial I complex in the parieto-occipital cortex of patients with BD (Ben-Shachar and Karry, 2008). A possible explanation for these results would be that increased mitochondrial complex activity occurs as a compensatory effect for ATP reduction observed in BD.
Results showed that Li was able to revert the increased activity of mitochondrial complexes I, II, II-III, but not the increase of IV complex activity. This may have occurred due to the fact that Li can act in a specific way in each brain region (Valvassori et al., 2010), and the response can be differentiated mainly as a function of the type of animal model induced (Frey et al., 2006), in this case, an animal model of the glutamatergic type (Ghedim et al., 2012).
These results suggest that this animal model of mania induced by Ket may have good face validity (induces hyperlocomotion in animals), predictive (Li reverses these effects), but may fail or have a limitation as to construct validity, since it does not the role of energy metabolism in the pathophysiology of BD is very well elucidated. Another study by Venâncio et al. (2015) showed that Ket increases mitochondrial nitric oxide synthase (mitNOS) by interfering with the function of mitochondrial complex I, thereby increasing NO and superoxide anion (O2.−) levels. Ket also increases apoptosis and ROS production in human neurons (Bai et al., 2013). An important factor in the pathogenesis of BD is a disorder of the glutamatergic system, and Li, as a mood stabilizer, can prevent behavioral and biochemical changes caused by KET in animals (Gazal et al. 2014; Saljoughi et al., 2023).
It is important to emphasize the vantage of this animal model in relation to other models of mania, which is a reproduction of glutamatergic changes induced by Ket, such as those found in the phase of mania in BD. In the post-mortem brains of patients with BD, there was an increase in the glutamate/glutamine ratios and a decrease in the levels of N-methyl-D-aspartate (NMDA) receptor subunits (Hashimoto et al., 2007; Rao et al., 2012). In addition, a study by (Dickerson et al., 2012) demonstrated increased levels of antibodies to a subunit of NR2 (NMDA receptor subunits) during an acute phase of patient mania. Therefore, this model may be more relevant to mimic a glutamatergic alteration than a change in energy metabolism (Saljoughi et al., 2023).
In conclusion, our study shows the increase in mitochondrial respiratory capacity in a glutamatergic animal model of mania. Therefore, we suggest an interaction between glutamate, which is involved in the pathophysiology of BD, and mitochondrial dysfunction. We have also shown that Li can reverse Ket-induced mitochondrial dysfunction. However, the protective effect of Li was not observed in all respiratory chain complexes. Future studies may better elucidate the relationship between treatment with Li and the relationship with mitochondrial and glutamatergic dysfunction in animal models of BD using Ket.
{kind=link}