Prevalence of Cryptosporidium spp.
The overall prevalence of Cryptosporidium in neonatal dairy calves was 48.7% (185/380, 95% CI: 43.5-53.8%), with 48.6% (108/222, 95% CI: 41.9-55.5 %) in Alaer and 48.7% (77/158, 95% CI: 40.6-56.8%) in Wensu. In Alaer, Cryptosporidium was detected most frequently in summer (53.7%, 29/54, 95% CI: 39.5-67.9%), followed by spring (44.9%, 22/49, 95% CI: 30.0-59.9%), winter (52.5%, 31/59, 95% CI: 39.0-66.1%), and autumn (43.3%, 26/60, 95% CI: 30.0-56.7%), while the prevalence of Cryptosporidium did not differ significantly among the seasons (χ2 = 1.865, df = 3, P > 0.05) (Table 1). In Wensu, the highest prevalence occurred in summer (61.0%, 25/41, 95% CI: 44.8-77.1%), then spring (56.4%, 22/39, 95% CI: 39.6-73.3%), autumn (39.5%, 17/43, 95% CI: 23.8-55.3%), and winter (37.1%, 13/35, 95% CI: 19.7-54.6%) (χ2 = 6.718, df = 3, P > 0.05) (Table 1, Fig. 1).
The prevalence of Cryptosporidium in neonatal dairy calves with and without diarrhea were 72.4% (113/156, 95% CI: 65.1-79.8%) and 32.1% (72/224, 95% CI: 25.8-38.5%), respectively, which differed significantly and there was a significant association between the occurrence of diarrhea and Cryptosporidium infection (χ2 = 59.760, df = 1, P < 0.01) (Table 2). In Alaer, the prevalence of Cryptosporidium with diarrhea in autumn (66.7%, 14/21, 95% CI: 44.1-89.2%) and summer (82.6%, 19/23, 95% CI: 65.0-100%) were significantly higher than those without diarrhea in autumn (30.8%, 12/39, 95% CI: 15.0-46.5%) (χ2 = 7.163, df = 1, P < 0.01) and summer (32.3%, 10/31, 95% CI: 14.2-50.3%) (χ2 = 13.463, df = 1, P < 0.01), respectively. However, the prevalence with diarrhea in winter 63.2% (12/19, 95% CI: 38.8-87.5%) and spring 58.8% (10/17, 95% CI: (32.5-85.2%) differed not significantly without diarrhea in winter (47.5%, 19/40, 95% CI: 30.8-64.2%) (χ2 = 1.267, df = 1, P > 0.05) and spring (37.5%, 12/32, 95% CI: 19.2-55.8%) (χ2 = 2.040, df = 1, P > 0.05), respectively. In Wensu, the prevalence of Cryptosporidium in neonatal dairy calves with diarrhea in the four seasons were 65.0% (13/20, 95% CI: 68.4-100%) in autumn, 75.0% (12/16, 95% CI: 50.7-99.3%) in winter, 85.7% (18/21, 95% CI: 44.1-89.2%) in spring, and 78.9% (15/19, 95% CI: 58.0-99.9%) in summer, which were significantly higher than those in calves without diarrhea in autumn (17.4%, 4/23, 95% CI: 0-35.1%) (χ2 = 10.143, df = 1, P < 0.01), winter (5.3%, 1/19, 95% CI: 0-17.9%) (χ2 = 18.093, df = 1, P < 0.01), spring (22.2%, 4/18, 95% CI: 2.4-44.2%) (χ2 = 15.890, df = 1, P < 0.01), and summer (45.5%, 10/22, 95% CI: 22.4-68.5%) (χ2 = 4.806, df = 1, P < 0.05), respectively (Table 2, Fig. 2).
Distribution of Cryptosporidium species
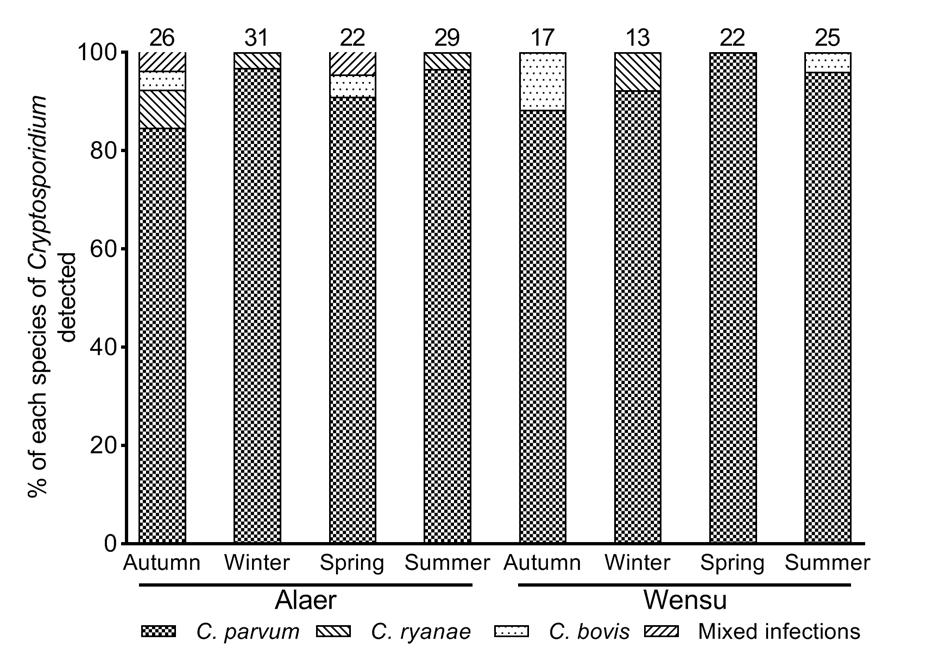
Three Cryptosporidium species, C. parvum (n = 173), C. bovis (n = 7), and C. ryanae (n = 3), and co-infections of C. parvum, C. bovis, and C. ryanae (n = 2) were detected with an RFLP analysis of the positive PCR products (χ2 = 367.600, df = 2, P < 0.01). C. parvum was clearly predominant in every season, whereas C. bovis and C. ryanae were found only occasionally, and co-infections were only seen in Alaer. Among the species detected, C. parvum (175/189) was the commonest species on the farms in Alaer (102/112) and Wensu (73/77) (Table 1).
In Alaer, the proportions of Cryptosporidium species identified in the four seasons were C. parvum 84.6% (22/26), C. bovis 7.7% (2/26), C. ryanae 3.8% (1/26), and co-infections 3.8% (1/26) in autumn; C. parvum 96.8% (30/31) and C. ryanae 3.2% (1/31) in winter; C. parvum 90.9% (20/22), C. bovis 4.5% (1/22), and co-infections 4.5% (1/22) in spring; and C. parvum 96.6% (28/29) and C. bovis 3.4% (1/29) in summer. In Wensu, the Cryptosporidium species distributions in the four seasons were C. parvum 88.2% (15/17) and C. bovis 11.8% (2/17) in autumn; C. parvum 92.3% (12/13) and C. ryanae 7.7% (1/13) in winter; C. parvum 100% (22/22) in spring; and C. parvum 96.0% (24/25) and C. bovis 4.0% (1/25) in summer (Fig. 3).
Subtypes of C. parvum
Among the 175 C. parvum-positive samples (102 from Alaer and 73 from Wensu), 173 were successfully sequenced and identified as two subtypes, IIdA14G1 (n = 94) and IIdA15G1 (n = 78) (χ2 = 2.960, df = 1, P > 0.05) (Table 2). In Alaer, the dominant subtype IIdA14G1 (n = 94, χ2 = 149.881, df = 1, P < 0.01) was identified in every season, and the difference in the IIdA14G1 detection rates in dairy calves with diarrhea (51.5%, 52/101, 95% CI: 41.2-61.7%) and those without diarrhea (41.6%, 42/101, 95% CI: 31.5-51.7%) was not significant (χ2 = 2.128, df = 1, P > 0.05). Interestingly, IIdA15G1 (n = 7) was only seen in calves without diarrhea. The subtype distributions in the four seasons were IIdA14G1 95.7% (22/23, 95% CI: 85.1-100%) and IIdA15G1 4.3% (1/23, 95% CI: 0-14.9%) in autumn; IIdA14G1 89.7% (26/29, 95% CI: 76.9-100%) and IIdA15G1 10.3% (3/29, 95% CI: 0-23.2%) in winter; IIdA14G1 85.7% (18/21, 95% CI: 68.4-100%) and IIdA15G1 14.3% (3/21, 95% CI: 0-31.6%) in spring; and IIdA14G1 100% (28/28) in summer (Fig. 4). In Wensu, C. parvum was not detected in dairy calves without diarrhea in winter, and IIdA15G1 was the only subtype identified. The IIdA15G1 detection rate was significantly higher in calves with diarrhea (80.3%, 57/71, 95% CI: 70.3-90.2%) than in calves without diarrhea (19.7%, 14/71, 95% CI: 9.8-29.7%) (χ2 = 52.085, df = 1, P < 0.01) (Table 2).
{kind=link}