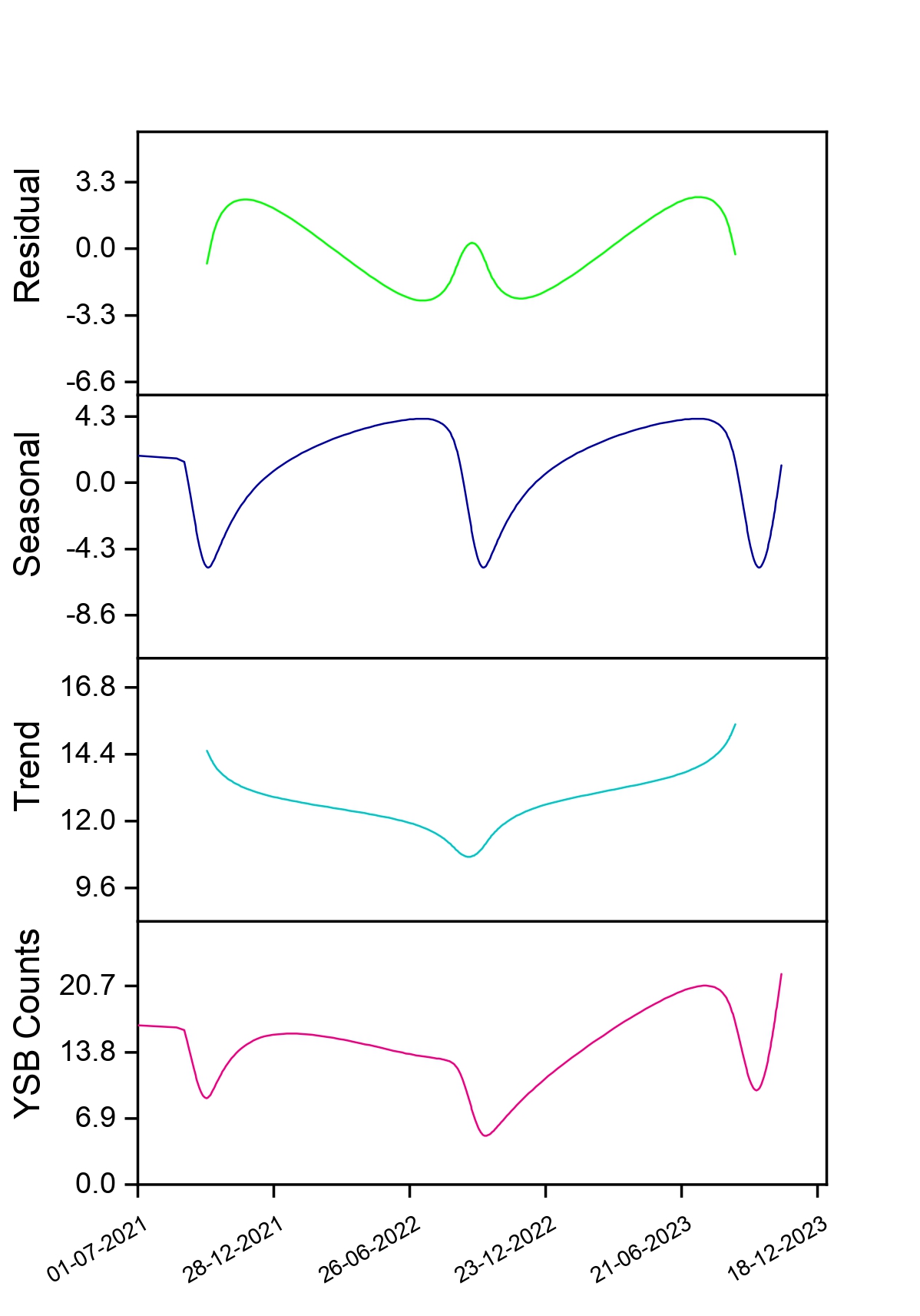
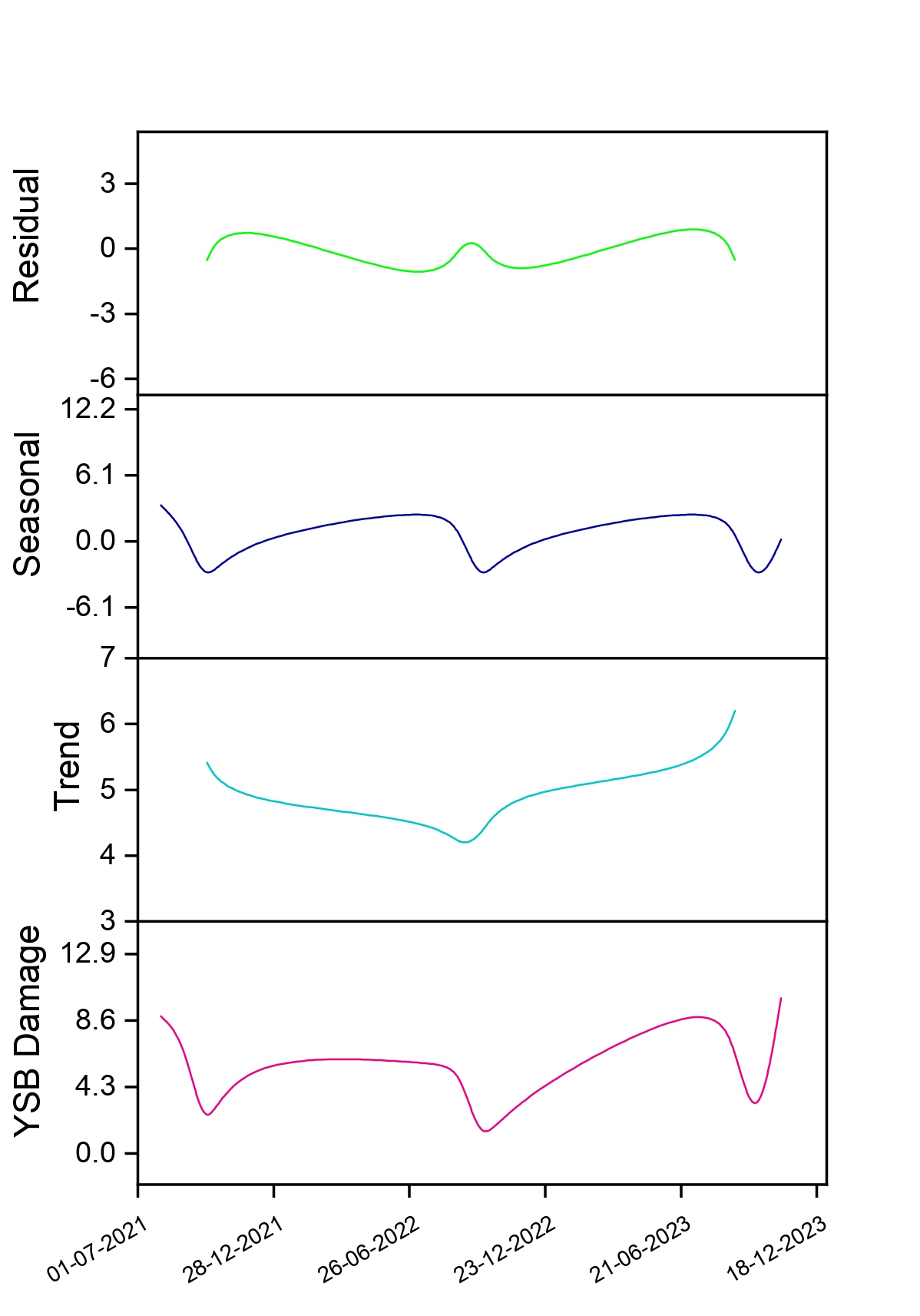
Understanding the factors affecting the spatial distribution/pattern of YSB in fragmented rice landscapes is an essential component of population ecology. It is vital for predicting YSB populations, enhancing sampling techniques and applying this knowledge to decision-making in pest management (Haila et al., 2002; Binns et al., 2000; Yang et al., 2018; Ribeiro et al., 2021). In this study, YSB moth populations and damage followed a ‘peak-fall-rise’ pattern in rice crops during the kharif season across the sampling locations (Fig. 4). An intraseasonal rise and fall of YSB populations and damage in rice have been previously reported (Kumar, 1995; Justin and Preetha, 2013; Patel and Singh, 2017; Baskaran, 2019; Jasrotia et al., 2019; Bapatla et al., 2024). Pathak (1968) opined that peaks in YSB populations reflect major seasonal effects, which have been misinterpreted as different broods. The principal factors -abiotic (surface air temperature, rainfall, day length, and humidity) and host plant phenology (Pathak, 1968) - define YSB population dynamics and damage in the study area. The occurrence of a warm and humid climate (Pathak, 1968; Adiroubane and Raja, 2010) and the excessive application of nitrogen fertilizer (Hirano, 1964; Singh and Sarkar, 2021) encouraged YSB populations and damage during the vegetative stage of the crop in August and September. Subsequently, the initiation of low surface air temperatures around 28°C (Kiritani and Iwao, 1967) and short-day lengths of 8–14 hours (Inoue and Kamano, 1957) in October induced diapause in YSB. In the last week of October, the availability of the susceptible stage of rice (panicle emergence) and rainfall terminated the diapause (Van der Laan, 1959), resulting in a buildup of YSB populations and damage in November. Our findings are corroborated by earlier studies on peak activity periods of YSB populations and damage during July, August and September (Varma et al., 2000), August and September (Justin and Preetha, 2013; Jasrotia et al., 2019), and September (Patel and Singh, 2017) in the kharif season.
A strong and positive association between surface air temperature and relative humidity with YSB populations and damage was proven in this study. This is further substantiated by the observation that high temperatures coupled with high relative humidity (Kumar and Sudhakar, 2001; Mishra et al., 2005; Hugar et al., 2010) in August and September, and low temperatures in November (Rahaman et al., 2014), favoured YSB population buildups and led to severe damage in rice. Although no previous studies have explained the relationship between LST and NDVI values with YSB populations or damage in rice, our study attempted to do so. We observed a statistically negative relationship between them, but the factors responsible for this association are not fully understood yet, and potential causes should be explored in future studies.
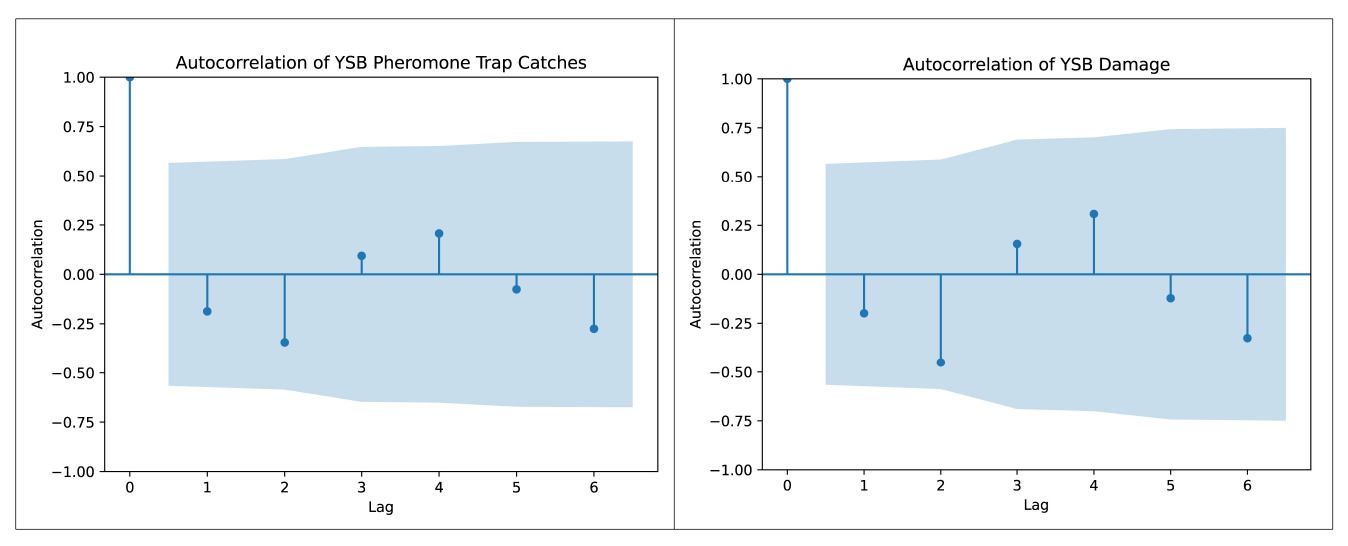
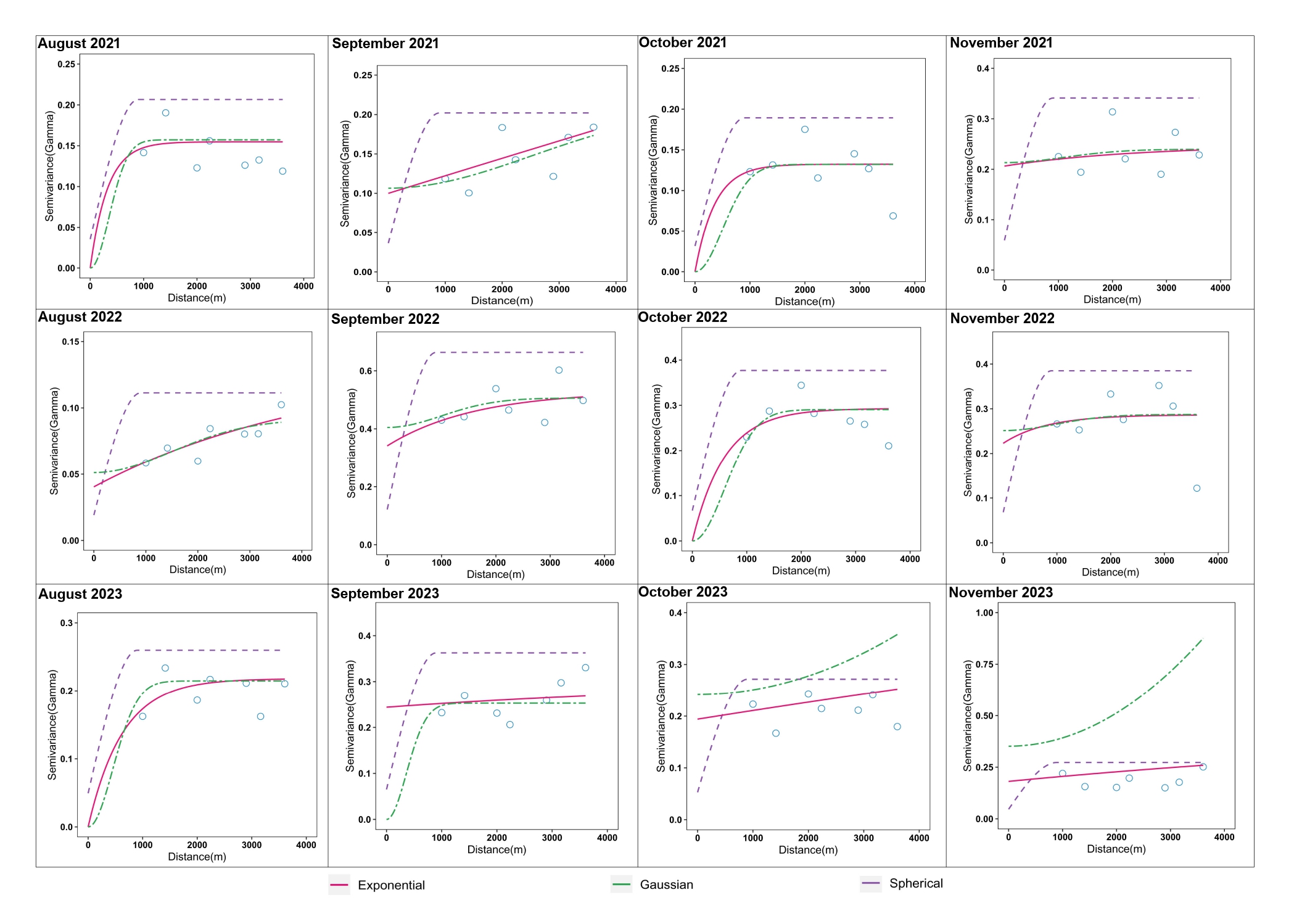
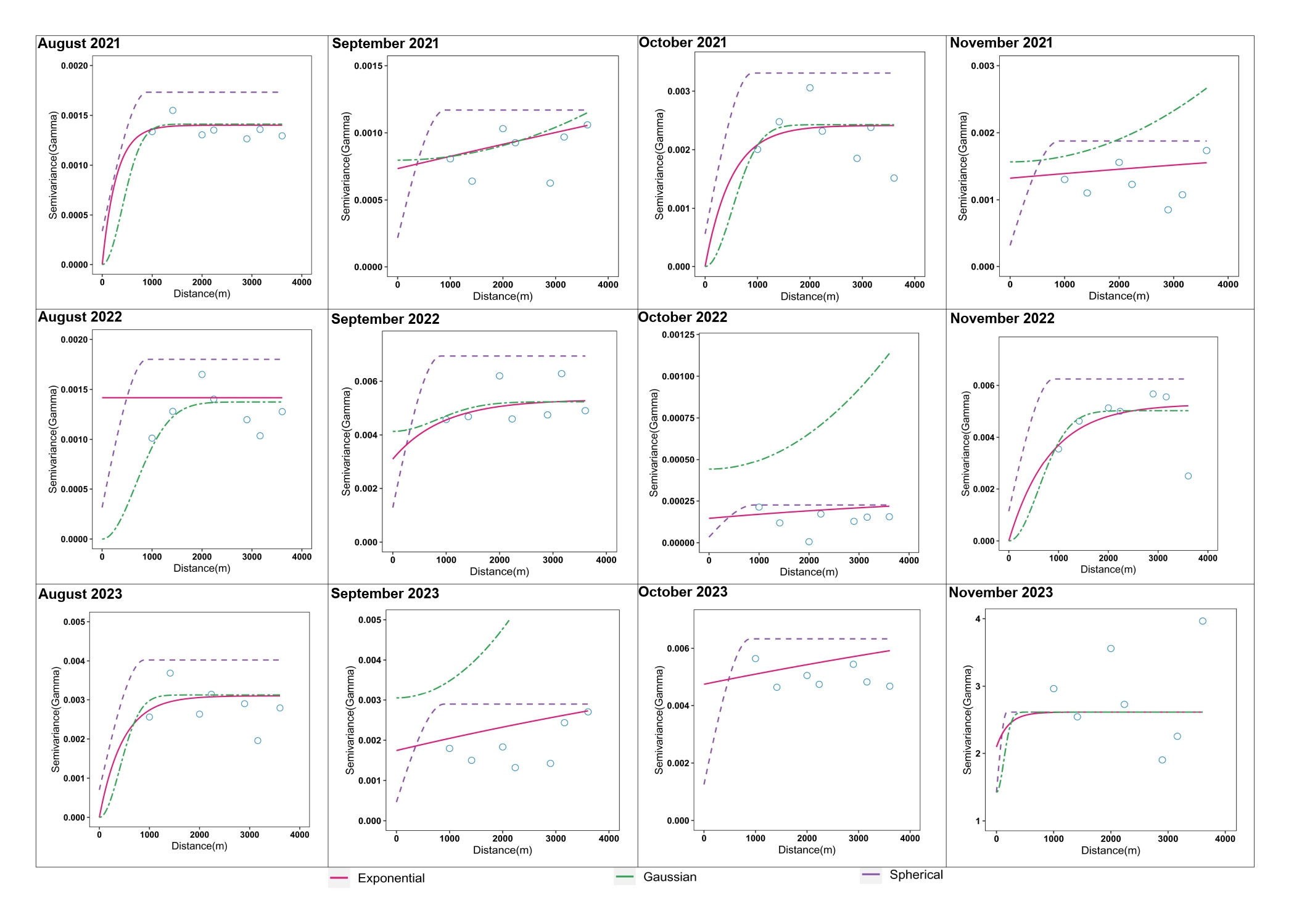
In addition to biotic, abiotic and host factors, a spatial dimension was introduced in this study to characterize YSB populations and their damage in rice. Temporal variations in YSB in rice have been reported using pheromone and light trap catches (Kumar, 1995; Baskaran et al., 2019; Gangappa et al., 2023; Bapatla et al., 2024) and damage assessments (Justin and Preetha, 2013; Patel and Singh, 2017; Jasrotia et al., 2019; Sharma and Sharma, 2023), but spatial points were not considered in earlier studies. To understand the effect of space on YSB, a semivariogram was developed through geostatistical analysis. Spatial aggregation of YSB populations and damage was detected using the semivariogram in all 12 sampling months, indicating weak to strong temporal aggregation across the rice fields in the study area (Suppl. Figure 4 and Suppl. Figure 5). Based on the spatial variance ratio, strong spatial aggregation/dependence was detected in four and five sampling months for YSB populations and damage, respectively. Although geostatistical tools have been utilized to understand the spatiotemporal distribution of several agricultural pests such as Helicoverpa armigera in pigeonpea (Seethalam et al., 2021), Plutella xylostella in crucifers (Li et al., 2021), Dalbulus maidis in corn (Ribeiro et al., 2021) and Chrysolina aeruginosa in desert vegetation (He et al., 2023), this study is the first of its kind to document the spatial distribution of YSB populations and their damage in rice.
In randomly sampled rice fields, visual-counting methods like pheromone trap catches and percent-damaged tillers (deadhearts and whiteheads) are the most common methods for YSB population monitoring and damage assessment (Jasrotia et al., 2019; Varma et al., 2000). However, these methods are laborious and time-consuming, especially in larger rice fields. Therefore, to develop reliable sampling methods for insect pest monitoring and population assessment, variogram models (spherical, Gaussian, and exponential) were used to estimate the range values that specify the aggregated distribution pattern (Shrestha et al., 2020; Seethalam et al., 2021). The range values also signify the end of spatial autocorrelation of YSB populations and damage in the space dimension (experimental area). Our findings suggest that an average sampling distance (range value) of 584.1 m and 673.33 m should be followed to reduce scouting fatigue for counting YSB pheromone trap catches and percent damage in large rice fields for monitoring purpose. These results may improve scouting and monitoring of YSB populations and damage in rice.
The results of Moran’s I indicate significant spatial clustering of YSB, suggesting that the population and damage are not randomly distributed but exhibit spatial dependence, which aligns with similar findings in spatial ecology (Li et al., 2024). The identification of YSB population/damage hotspots using the Getis-Ord Gi* statistic further emphasizes that the southeast direction of the sampling area (L27, L28, L29, L30, L31, and L32) had more hotspots, which may be influenced by natural and socio-economic factors (Li et al., 2024). These findings are crucial for targeted pest control strategies, as focusing on identified hotspots can optimize resource allocation and enhance control efficiency. The propensity of insect movement in patches and settlement in space in a contagious fashion, within or between habitats, tends to be directional and not random (Stinner et al., 1983). This spatial pattern of immigration of YSB, prior to colonizing and increasing in population size, can be managed by following “precision targeting” the hotspots identified in this study. Thus, one of the possible implications of this study for managing YSB is that the early installation of a reasonable number of sex pheromone traps, along with the placement of bioagent trichocards(containing Trichogramma japonicum parasitized eggs of Corcyra cephalonica) in hotspot locations, can further prevent the movement of YSB. In this way, the use of pesticides can be minimized.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}