Defined by lack of T cell infiltration within tumor, non-T-cell-inflamed microenvironment, was one of the major cause for immunotherapy resistance. Our study collected evidence that PTEN loss was one of the tumor-intrinsic mechanism contributing to an immune exclusion phenotype. We profiled the correlation of PTEN loss with T-cell-exclusion microenvironment across different solid tumors based on TCGA database, and found that both genomic loss (loss-of-function mutation or copy number deletion) and low expression of PTEN were associated with immune exclusion in a pan-cancer setting. PTEN’s impact on tumor immune microenvironment might be mediated by PI3K pathway, whose activation was also associated immunosuppressive phenotype in multiple solid tumors. These findings not only facilitate our understanding in the correlation between PTEN loss and immunotherapy resistance, but also inspire continued pursuit on immune modulating strategy to augment immunotherapy.
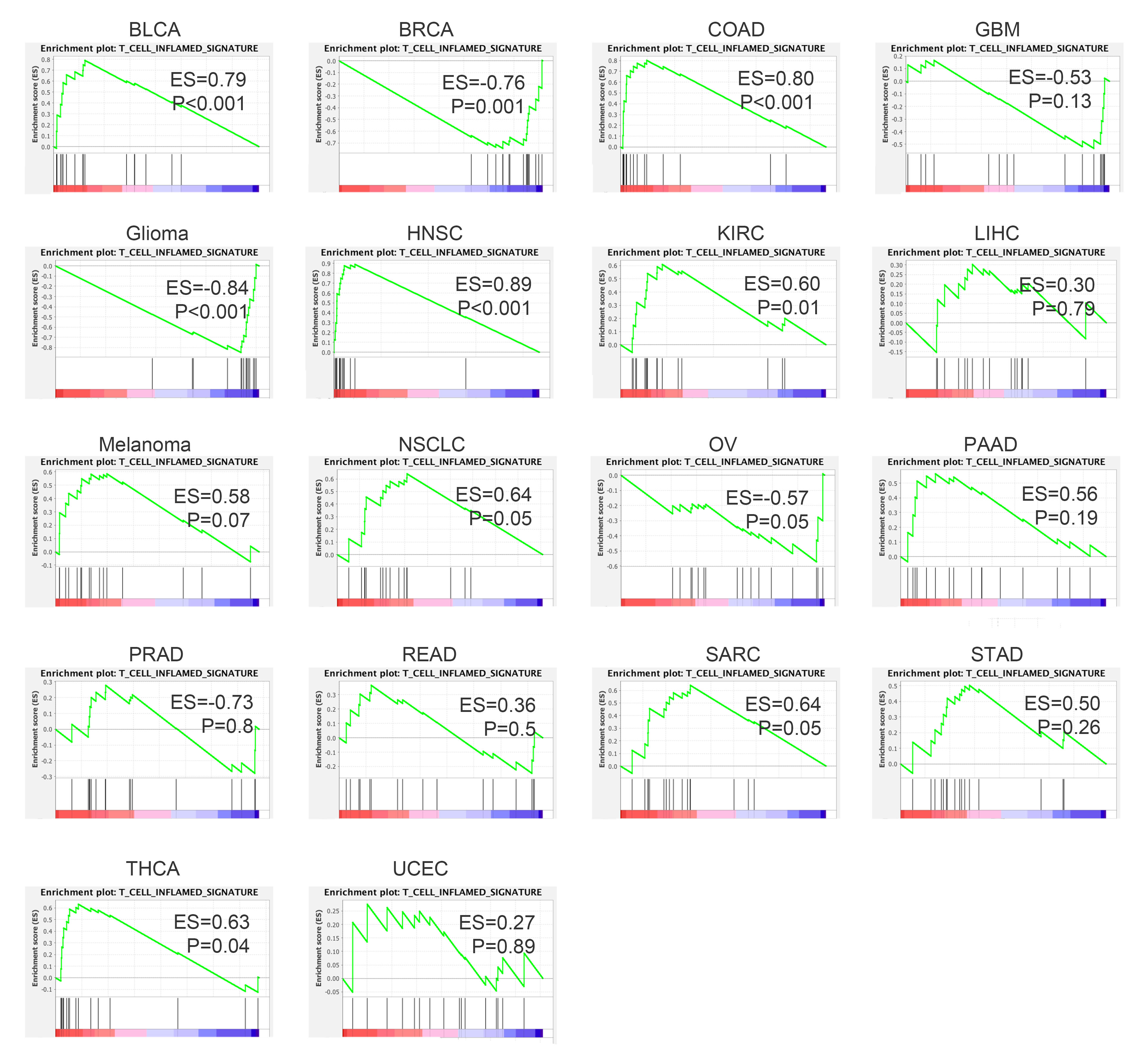
In this study, we evaluated the correlation of genomic alteration and expression of PTEN with tumor T cells infiltration. Consistent with previous findings, we found both genomic loss or reduced expression of PTEN were associated with decreased T cell trafficking and immune suppressive microenvironment in board range of malignancies[8, 9, 19, 30]. What differs our findings from the published studies is that we first reported T cell infiltration level was positively correlated with PTEN expression level. Also, reduced expression in PTEN predicted poor response to immunotherapy and worse outcome. These findings indicated that PTEN’s impact on immune microenvironment is expression dependent. A recent published study indicated that overexpression of PTEN in tumor cells can enhance T-cell mediated tumor clearance, which also proved that it’s the expression level rather than functional status of PTEN that matters[31]. Another interesting finding in the present study is the strong correlation between PTEN expression and memory T cells (central memory T cells and effector memory T cells). Studies on TME had revealed that memory T cells were mainly located in tertiary lymphoid structures (TLS), which encompass abundant immature T cells, B cells and play an important role in tumor cell clearance[32]. The presence of TLS is not only associated with improved clinical outcome in multiple cancers, but also found to promote immunotherapy response in patients with melanoma or renal cell carcinoma[33, 34]. It’s possible that the involvement of PTEN loss in immunotherapy resistance is partial mediated by TLS. If it’s the case, it can explain the finding reported in a recent study that tumors with wide type PTEN manifested significant higher effector T cells as compared to tumors with mutant PTEN only after the treatment of immunotherapy, when memory T cells will be activated and differentiate into functional T cells [7].
Of note, the analysis of PTEN’s correlation with immune infiltration from the perspective of expression level and genomic alteration status didn’t yield exactly concordant results. For example, we didn’t find significant correlation of PTEN expression with the infiltrating level of CD8 T cell and Th2 cell, which though, were significant reduced and increased respectively in multiple tumors with genomic PTEN loss. Also, memory T infiltration was strongly correlated with PTEN expression, but only demonstrated significant reduction in selected tumor types with PTEN loss. These discrepancy may be attributed to the miscellaneous signal of PTEN that derived from stromal cells in addition to cancer cells. When immune cells or other stromal cell would also contribute to part of the expression level of PTEN, genomic data of PTEN (mutation and copy number variants) were more likely restricted to the tumor cells. Therefore, although we observed a positive correlation between PTEN expression and memory T cells infiltration, we can’t rule out the possibility that correlation is partially attributed to the high expression of PTEN in certain immature T cells. Another explanation for this discordance could be the way we defined loss-of-function mutation for PTEN. Loss-of-function mutations include mutations that cause inhibition of PTEN catalytic activity or mutations that lead to unstable truncated proteins. As the immune modulating function of PTEN might be independent of its catalytic activity, loss-of-function mutation is not necessary synonymous to loss of immune modulating function[35].
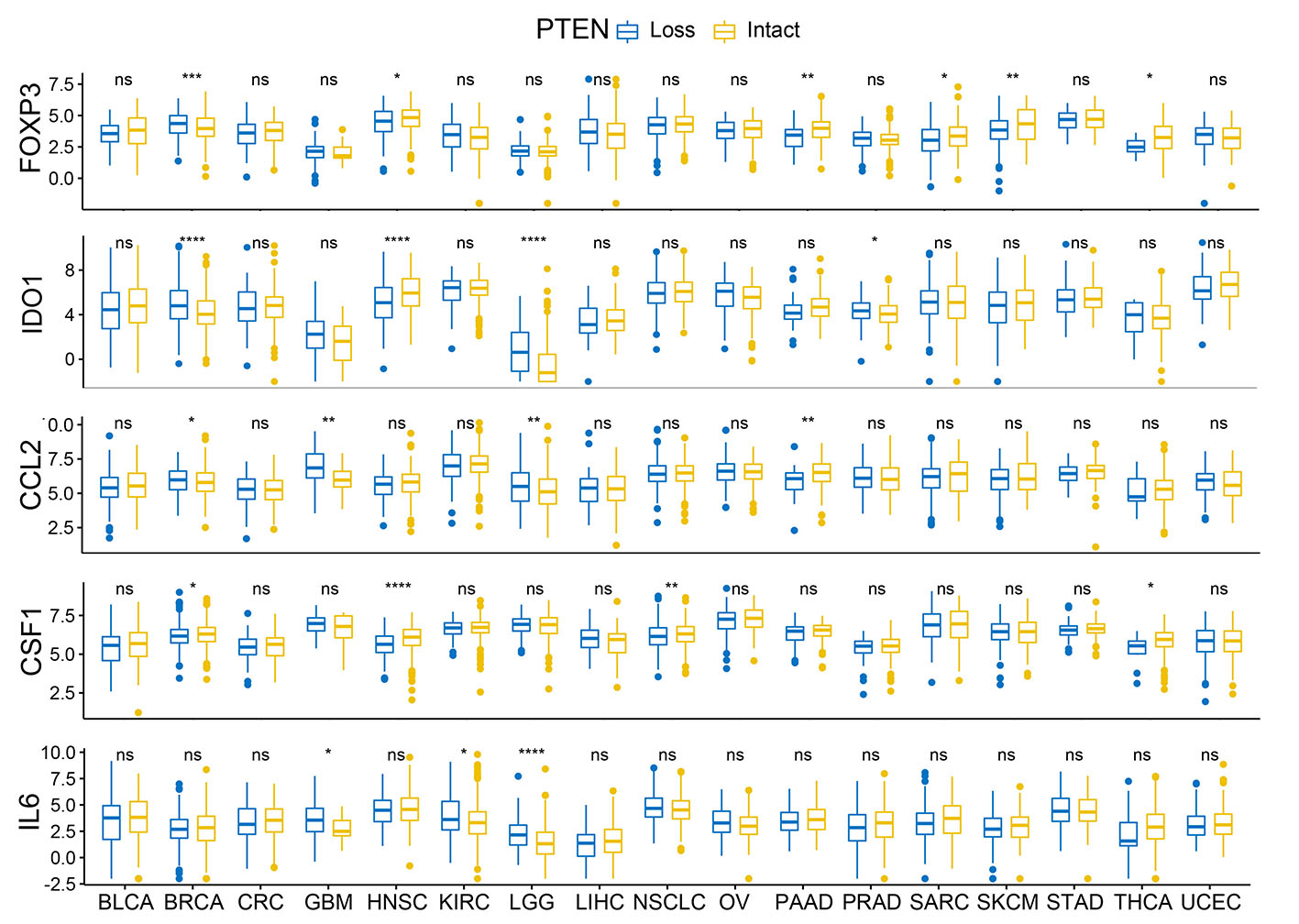
Existing study had shown that PTEN loss in tumor cells were associated with increased expression of immunosuppressive cytokines, especially VEGF, which reduce T trafficking as well as its cytotoxic function in TME [9]. Loss of PTEN in glioma was also reported to be associated with increased PD-L1 expression and immune escape[36]. There is still evidence indicates the correlation between PTEN loss on tumor cells and elevation of immunosuppressive markers like IDO1, FOXP3, CCL2, CSF1 ect.[37, 38]. Consistent with the published findings, we also found that PTEN loss is associated with increased mRNA expression of VEGFA, IDO1, IL6, CCL2, CSF1 in a wide spectrum of solid tumors. The expression level of PD-1, PD-L1 and FOXP3 were not altered by PTEN loss in most tumors, or even demonstrated reduced expression in some tumors, which might be attributed to the reduced T cells abundance associated with PTEN loss.
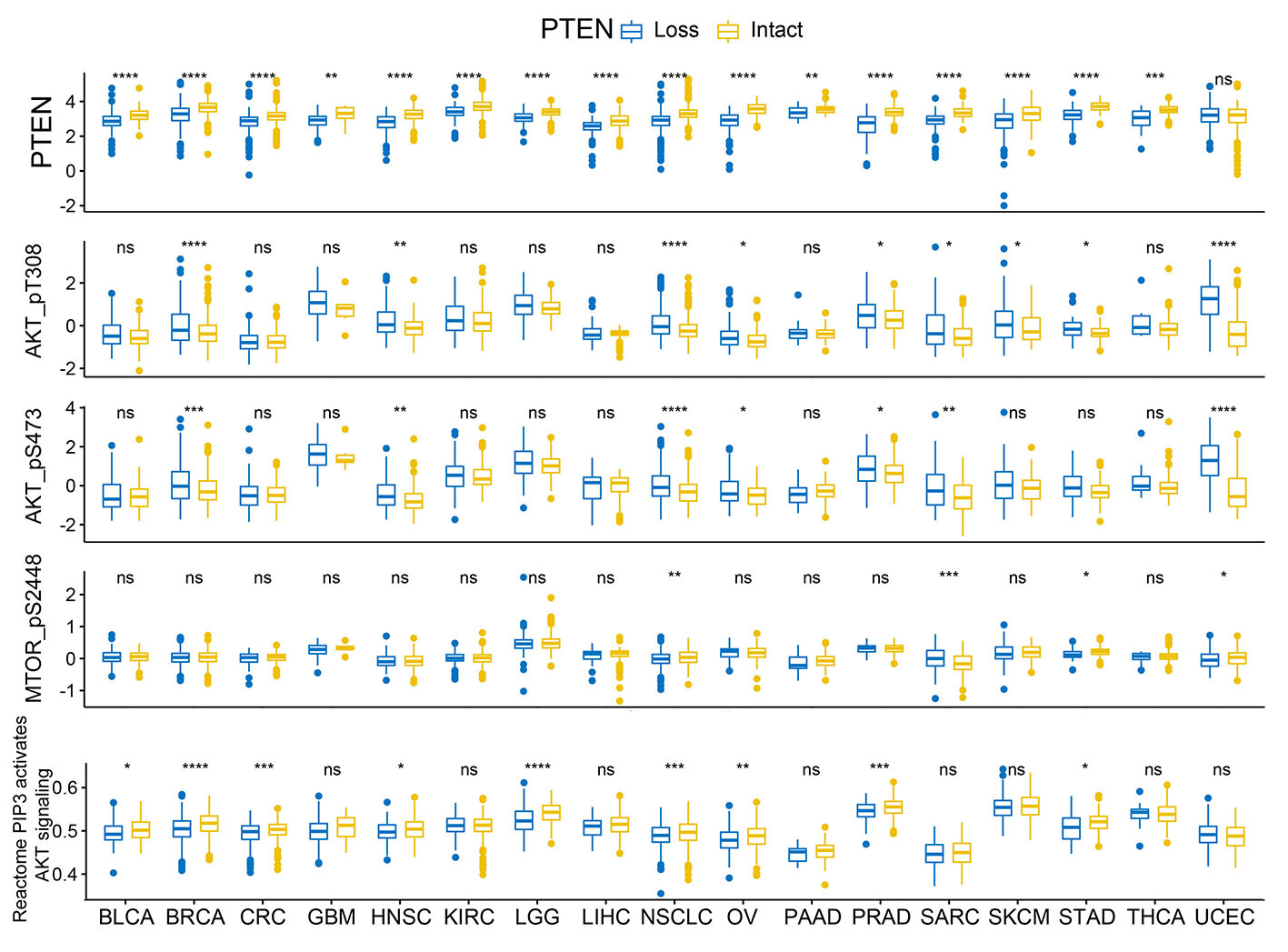
Our findings confirmed that PTEN loss in tumor cells was associated with T cell exclusion and an immunosuppressive microenvironment in solid tumors, though the underlying mechanism of which remains a puzzle. As we all know, the cornerstone discovery of PTEN’s biology was the negative regulation of the pro-oncogenic PI3K-AKT-mTOR pathway mediated by dephosphorylation of substrate PIP3[2, 27, 39, 40]. Quite a few studies had tried to tackle the immune modulating mechanism of PTEN from the perspective of PI3K-AKT-mTOR pathway alteration. James S. Waldron etc. found that PTEN loss associated with activation of the PI3K-Akt-mTOR pathway and led to a autologous T-cell apoptosis in glioblastoma, which can be diminished by treatment with inhibitors of PI3K-Akt-mTOR pathway[41]. Another study also indicated that PTEN-depleting melanoma cells promoted the expression of immunosuppressive cytokines in a PI3K-dependent manner[37]. We do observed a negative correlation of p-AKT level and PI3K pathway scoring with T cell infiltration in multiple tumor types, though the correlation is not as strong as we expected. It’s worth mentioned that the data regarding p-AKT level and PI3K pathway scoring were derived from tumor cells as well as the infiltrating immune cells, which can obscure the correlation between T cell filtration and PI3K pathway activation in tumor cells. What’s more, we observed similar changes on T cell infiltration in tumors with genomic gain in PIK3CA/PIK3CB as that in tumors with PTEN loss, which support the hypothesis that PI3K activation and PTEN loss yield similar impact on tumor immune microenvironment. All these findings indicated that the immune modulating effect of PTEN is at least partially attributed to PI3K pathway alteration, although some non-canonical functions of PTEN might also play a role amidst. As indicated in our data, PTEN loss is not synonymous with PI3K-Akt-mTOR activation, which is also revealed in published literature[42]. Except for the typical downregulation of PI3K-Akt-mTOR pathway, PTEN also exerts a series of non-canonical functions in a enzymatic or non-enzymatic manner[27, 35, 43, 44]. Emerging studies revealed that PTEN loss in prostate cancer resulted in an immunosuppressive tumor microenvironment through the activation of the Janus kinase 2 (JAK2)–signal transducer and activator of transcription 3 (STAT3) pathway and the subsequent secretion of immunosuppressive chemokines[38]. There are also studies reported that PTEN loss induced secretion of immunosuppressive cytokines via the activation of NF-κB pathway[37, 45]. The exact pathway by with PTEN exerts the immune modulating function is yet to be verified by further studies.
In addition to PTEN loss, more and more oncogenic events had been associated with tumor intrinsic resistance to immunotherapy. WNT/b-catenin pathway activation is also constantly detected in tumor developing resistance to immunotherapy[30, 46]. A recent published study has proven the correlation between wnt/b-catenin pathway activation and immune exclusion across human cancers[47]. Also, gain-of-function mutations in FGFR3, as well as activation of PPAR-γ pathway, have been associated with the T cell exclusion in tumor microenvironment in bladder cancer[18]. EGFR mutation in lung cancer is a well-known oncogene that impairs response to immunotherapy, which is attributed to an uninflamed phenotype and weak immunotherapy[48–50]. Of course, the above-mentioned oncogenic events didn’t account for all the tumors with the non-T-cell-inflamed phenotype, the full picture of which warrants continued effort on further research.
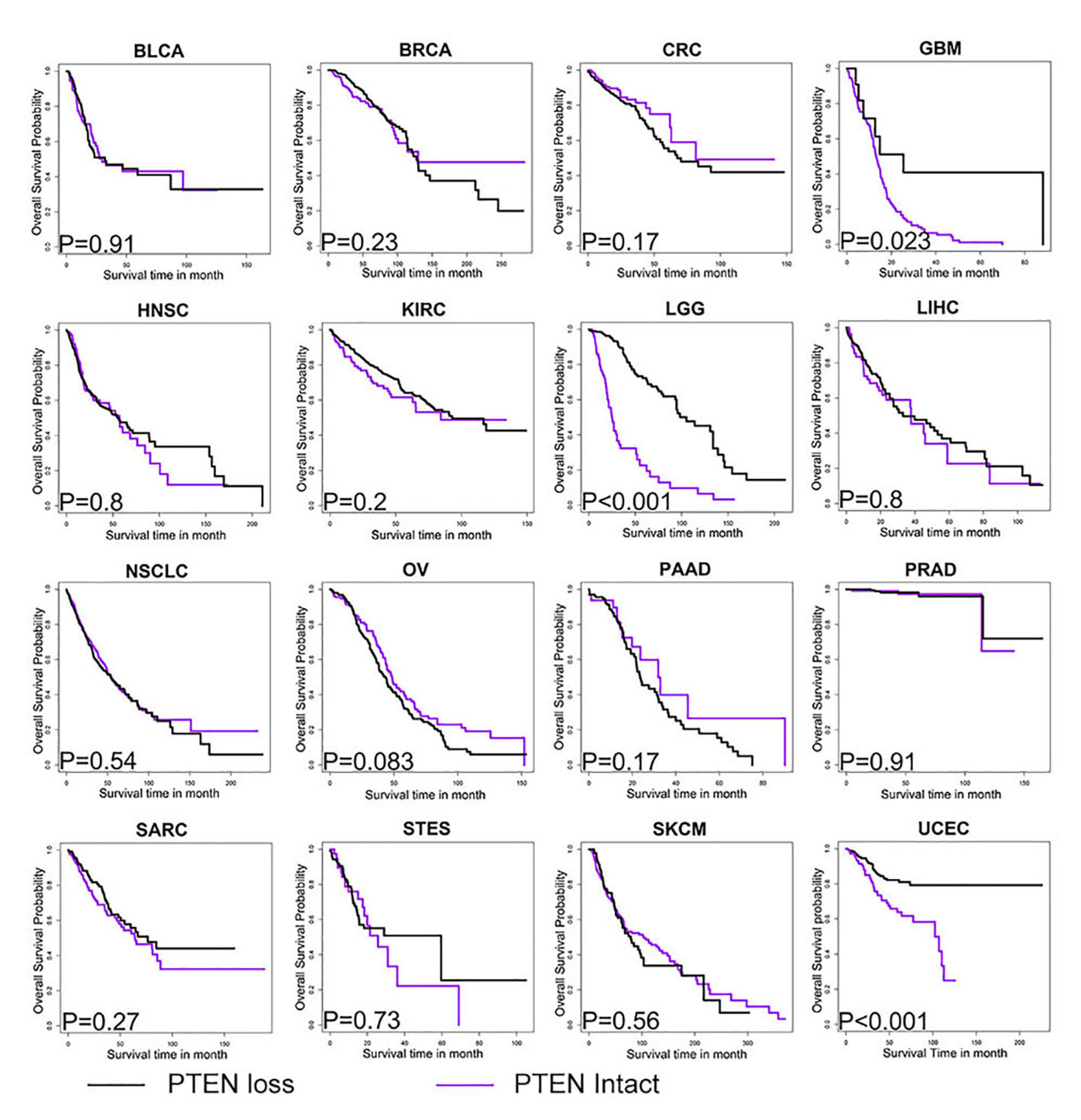
We found that PTEN status was a robust predictor for response to immunotherapy including PD-1 blockade and CTLA-4 blockade, although it was not associated with prognosis among patients without receiving immunotherapy. Our findings demonstrate that PTEN functions at the interface between cancer and tumor microenvironment and can eventually alter therapeutic outcome of immunotherapy. In the era of immunotherapy and precision medicine, our findings will translate into the molecular approach to classify the subpopulation of cancer patients that have greater chance of responding to immunotherapy. Also, with the implication on the mechanism by which PTEN modulate the immune microenvironment, we might be able to develop strategy to manipulate immune landscape and augment the therapeutic efficacy of immunotherapy in the clinical practice. Inhibitors targeting PI3K-Akt-mTOR pathway had shown promising efficacy in improving immunotherapy response in selected cancer type[9, 41, 51]. However, considering the multifaceted function of PTEN in all kinds of biological process, further study is warranted to bring in-depth understanding into its role in immune microenvironment and immunotherapy before we can actually exploit these knowledge in clinical practice.
Even with the promising findings, limitations of the present study need to be clearly addressed. First of all, the current work was merely based on the in silico analysis of TCGA, with all the immune parameters inferred from the transcriptome data. All these findings need to be confirmed by further study integrating IHC data or protein data. Also, data regarding PTEN status or PI3K-Akt-mTOR alteration were not derived from pure tumor cells, but from the tumor mass containing tumor cells as well as a small subset of stromal cells, infiltrating immune cells, whose confounding effect cannot be ruled out. Lastly, our analysis regarding genomic alteration of PTEN is limited to point mutations, small insertions and deletions, as well as copy number variants, whereas other types of genomic changes such as gene fusions or large-scale structural variants remain to be explored.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}