Despite the prevalence of gastric cancer, there is a current shortage of reliable biomarkers that can help predict prognosis and guide treatment decisions (15). Thus, it is crucial to find biomarkers that help with early detection and identify new treatment targets for GC.
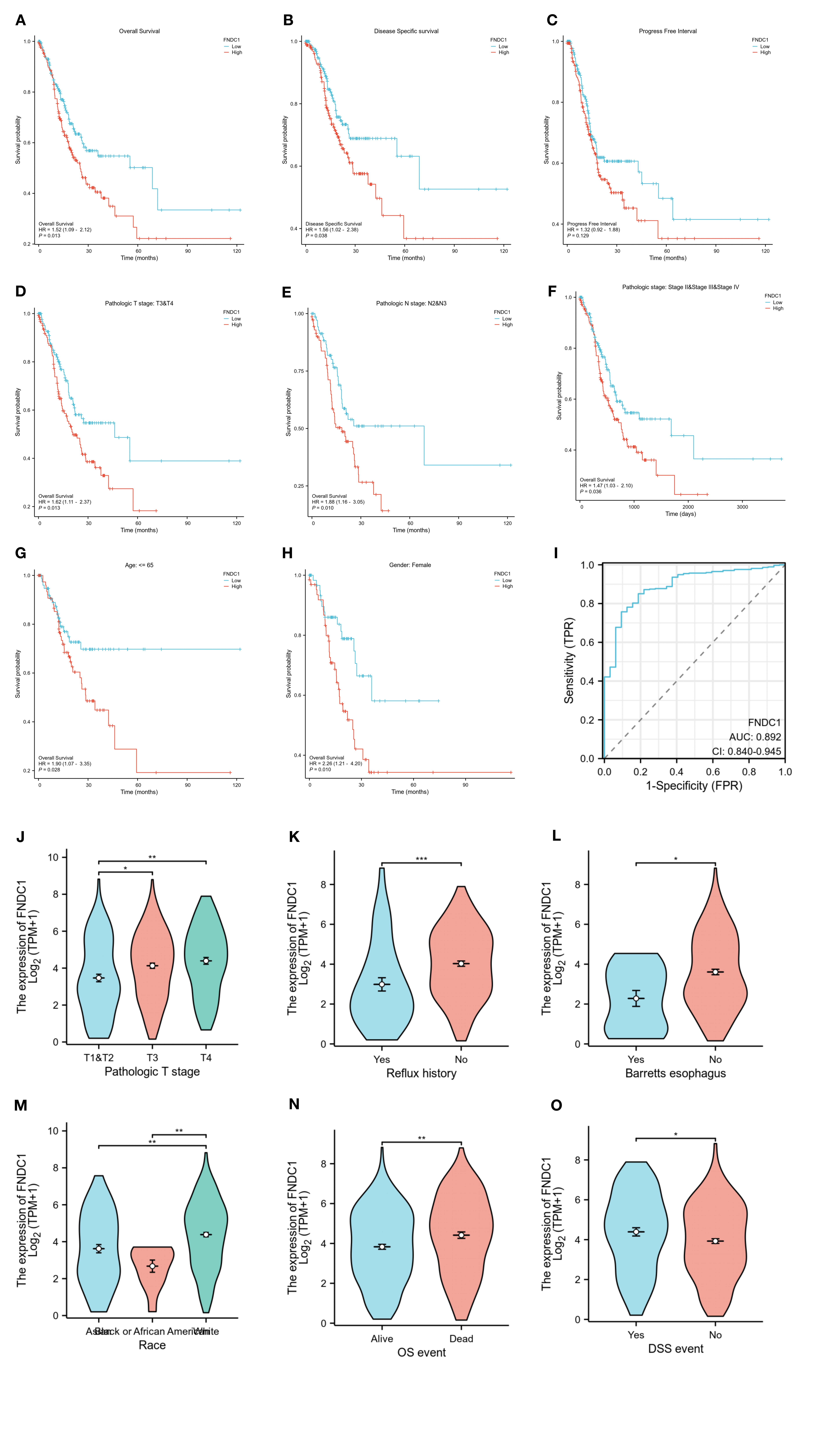
FNDC1 is an emerging biomarker associated with the development of various malignant tumors. Elevated levels of FNDC1 expression have been correlated with a poorer prognosis in gastric cancer patients (16, 17). This protein has a unique localization pattern, expressed in the nucleus and secreted into the extracellular matrix. FNDC1's dual localization gives it characteristics of both an intracellular protein and a secretory factor. This dual role implies that FNDC1 may be crucial in cellular functions and tumorigenesis. Therefore, FNDC1 has significant potential as a biomarker for cancer detection and could open new avenues for therapeutic intervention.
Reddy et al.(18) found that high levels of FNDC1 expression in head and neck squamous cell carcinoma significantly affect patient prognosis. At the same time, Chen et al. (19) discovered that FNDC1 is overexpressed in breast cancer and acts as an oncogene. Their study showed that suppressing FNDC1 inhibits the PI3K/Akt signaling pathway, which is essential for cancer cells' survival and proliferation. Moreover, other studies have identified FNDC1 as a potential prognostic indicator in breast cancer, highlighting its significance as a biomarker for clinical outcomes (20). Recent research (3) has linked FNDC1 to VEGF-regulated cellular activities such as angiogenesis, migration, and proliferation.
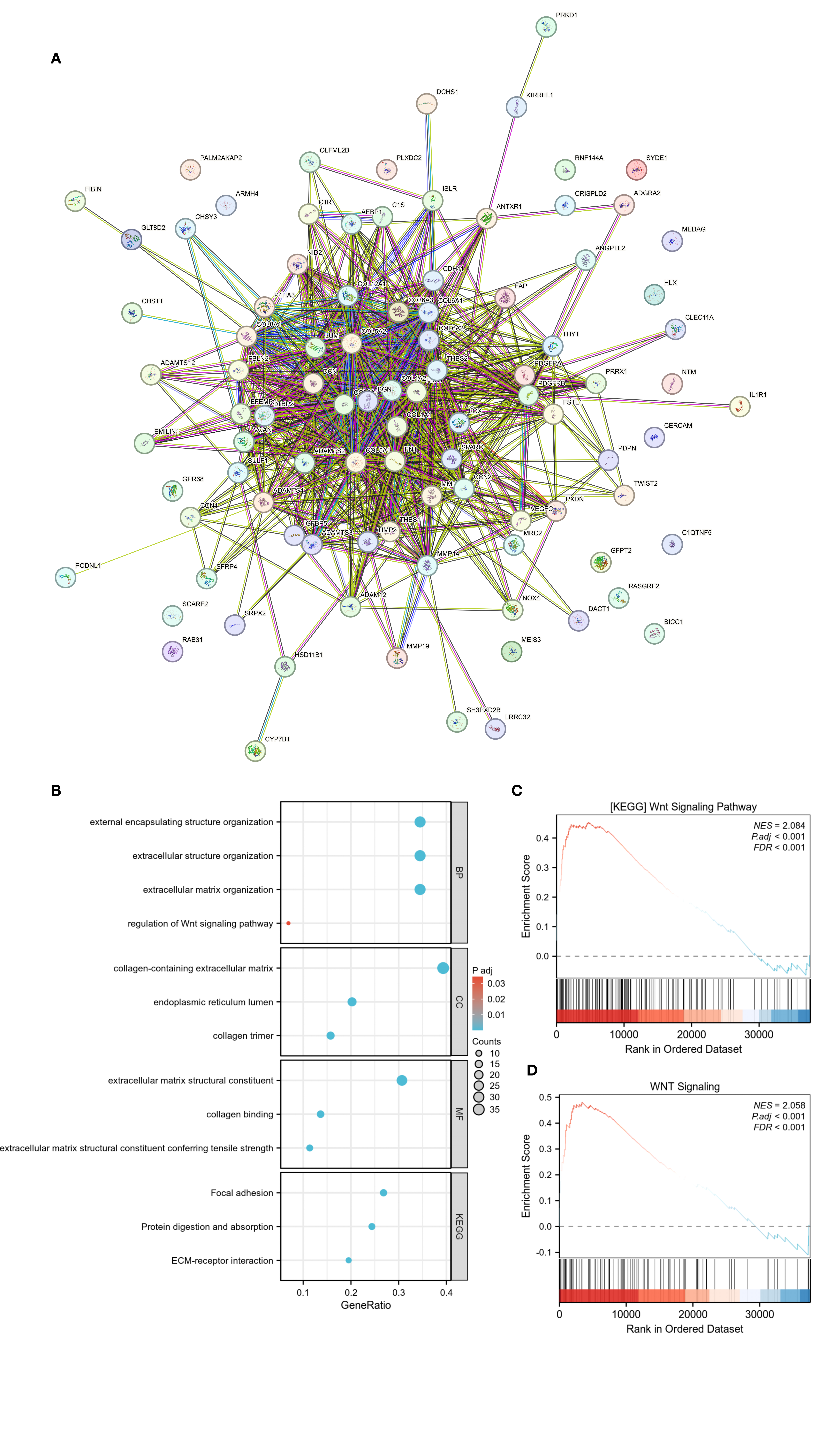
Additionally, microRNA-1207-3p induces apoptosis in prostate cancer cells by targeting FNDC1 (21). Recent studies indicate that FNDC1 is upregulated in GC, which promotes EMT and is linked to poor patient prognosis (16, 17, 22). These findings highlight FNDC1's complex role in cancer development and its potential as a prognostic biomarker in GC. Nevertheless, the molecular mechanisms by which FNDC1 facilitates EMT in GC cells remain largely unexplored. Our previous research demonstrated that FNDC1 promotes EMT by activating the Wnt/β-catenin signaling pathway in laboratory and animal models. Further delineation of these mechanisms is crucial for a thorough understanding of FNDC1's role in GC progression and for developing targeted therapeutic strategies.
This study aims to clarify how FNDC1 activates the Wnt/β-catenin signaling pathway. In this pathway, the stability of cytosolic β-catenin acts as a crucial regulatory switch. This stability is controlled by the Axin-GSK3β-APC complex, which is essential for regulating β-catenin levels and the activation of Wnt signaling. The expression levels of the Axin protein are recognized as a critical factor that depends on concentration within the β-catenin destruction complex (23). This indicates that both the expression levels and the functional integrity of Axin adjust the cellular response to Wnt signaling. The Axin family consists of two paralogs, Axin1 and Conductin/Axin2. Recent studies show Axin2 has a weaker suppressive effect on canonical Wnt signaling than Axin1(24).
Our investigation reveals that FNDC1 promotes the ubiquitination and degradation of Axin1 and disrupts its interaction with GSK3β and APC. The Wnt signaling cascade is activated when Wnt ligands bind to Frizzled and LRP5/6 receptors on the cell membrane, which triggers Dvl protein activation. This activation recruits Axin to the plasma membrane, which is targeted for ubiquitination and degradation, thus relieving its inhibition of β-catenin activation (12). The above process underscores the dependency of Axin ubiquitination on Dvl protein activation. We found that FNDC1 does not alter the expression levels of Frizzled1/2 and LRP5/6, but it significantly increases the protein levels of Dvl1/2/3. The proteasome inhibitor MG132 confirmed that FNDC1 affects Dvl1/2/3 at the post-transcriptional level, meaning it influences protein levels rather than gene expression.
Dishevelled (Dvl) is a pivotal intracellular signaling molecule that positively modulates the Wnt signaling pathway (25). The activation of Dvl is controlled by the Gβγ subunit of the heterotrimeric G protein in this pathway. The heterotrimeric G protein is a complex comprised of three distinct subunits: α, β, and γ. This complex is integral to cellular signaling. Precisely, the Gβγ subunits form a non-covalent dimer. This dimer is often considered functionally monomeric due to its strong association and resistance to dissociation under non-denaturing conditions (7). The βγ subunit complex primarily localizes to the inner side of the plasma membrane, where it interacts with downstream effector molecules, transducing signals from the cell surface to the intracellular environment. Extant research has identified the Gβγ subunit, a critical mediator in signal transduction, as playing a significant role in the pathogenesis of various malignant tumors, including breast and ovarian cancers (8). Notably, the study by Nathalie et al. (26) has highlighted the interplay between Gβγ and Dvl as crucial in modulating the suppression of the Wnt/β-catenin signaling pathway. During Wnt signaling activation, the binding of Gβγ to Dvl is instrumental in preventing the phosphorylation of β-catenin by the Axin complex, thereby facilitating β-catenin stabilization and averting its degradation. Previous research found that high levels of FNDC1 in AGS and SGC7901 cell lines form a complex with Gβγ and VEGFR2. This complex promotes VEGFR phosphorylation and activates the RAS-MAPK or PI3K-AKT signaling pathways (27).
We performed a series of co-IP experiments to validate the proposed molecular mechanism depicted. Our results show that FNDC1 regulates Dvl1 expression by interacting with Dvl1 and Gβ2. FNDC1 can competitively bind to Dvl1, preventing Dvl1 from binding to Gβ2 on the inside of the cell membrane. This reduces the affinity of the Gβγ subunit for Dvl1. As a result of this competitive binding, the Gβγ subunit dissociates from Dvl1. This leads to increased levels of Dvl1 inside the cell and promotes the degradation of Axin1. To determine the interaction between FNDC1 and Gβ2, we created a series of Gβ2 protein deletion mutants to identify the exact binding site. Following a systematic analysis, we localized the primary interaction domain to the amino acid sequence spanning positions 224 to 254. Our mutagenesis study confirmed that this specific region, crucial for mitochondrial fusion and cellular signaling, is the critical binding domain. Prior literature supports the biological relevance of this fragment in cellular processes (28, 29).
In conclusion, our study shows that FNDC1 positively influences the Wnt/β-catenin signaling pathway. This research improves our understanding of FNDC1's role in the β-catenin destruction complex. We have demonstrated that FNDC1 disrupts the interaction between Gβγ and Dvl by competing with Gβ2 in GC cells. This disruption stabilizes Dvl, which promotes the continuous activation of the Wnt/β-catenin pathway. As a result, it enhances GC cells' invasive and metastatic potential. Our findings indicate that FNDC1 is a new oncogenic factor in GC, suggesting it could be a potential therapeutic target.
{kind=link}
{kind=link}
{kind=link}