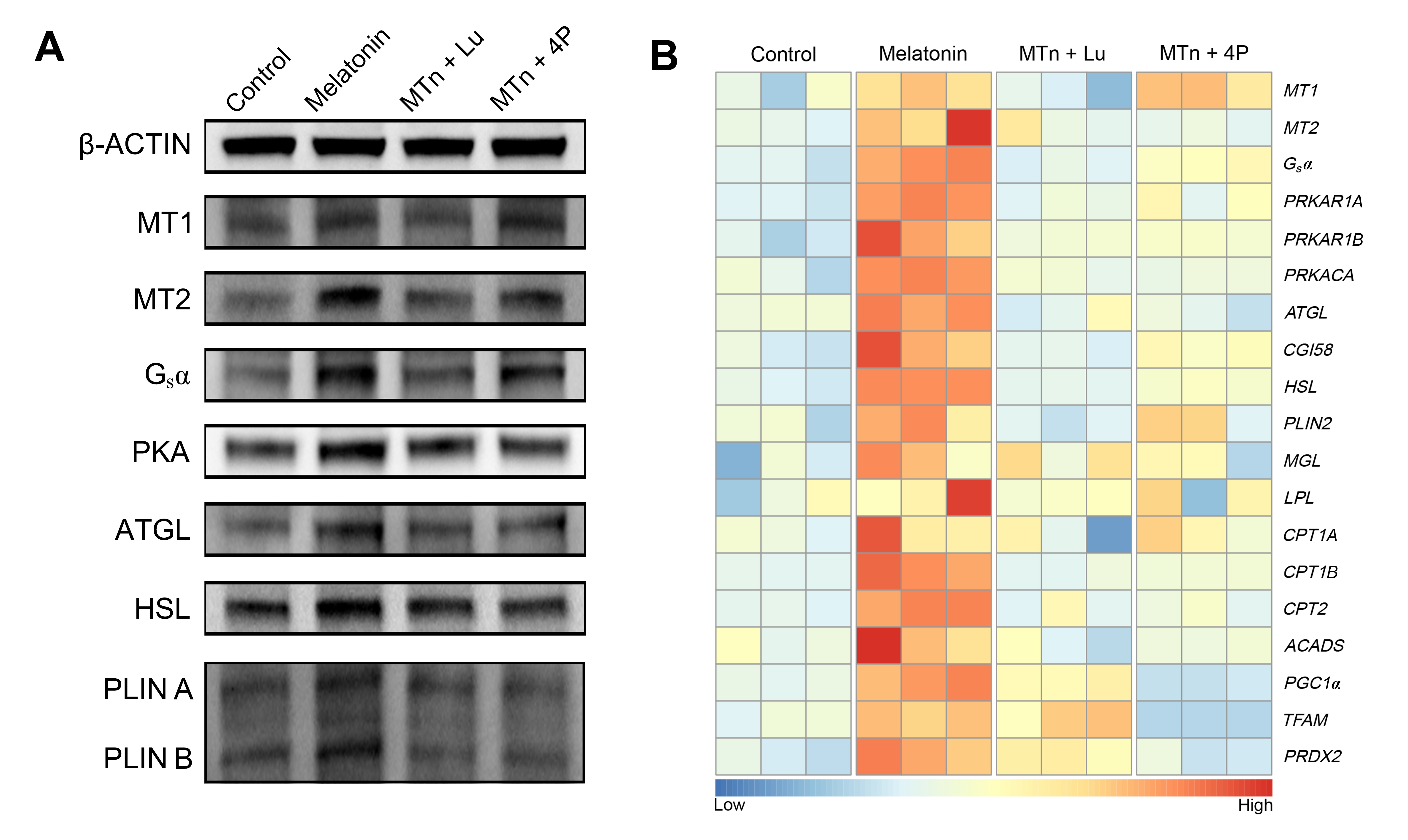
It is well known that G protein-coupled receptors (GPCRs) are the most functional cell surface proteins and mediate a series of physiological processes [26]. The melatonin receptors, MT1 and MT2, are GPCRs that receive external melatonin signals and are also involved in a vast array of physiological processes [27–29]. Nevertheless, the beneficial effects of melatonin on the reproductive system have been explained simply as being due to indirect, antioxidant effects, and so little information is available on the underlying mechanism by which melatonin concretely acts on porcine COC development. Therefore, a better understanding of the melatonin receptor-mediated intracellular signaling pathway in cumulus cells is required.
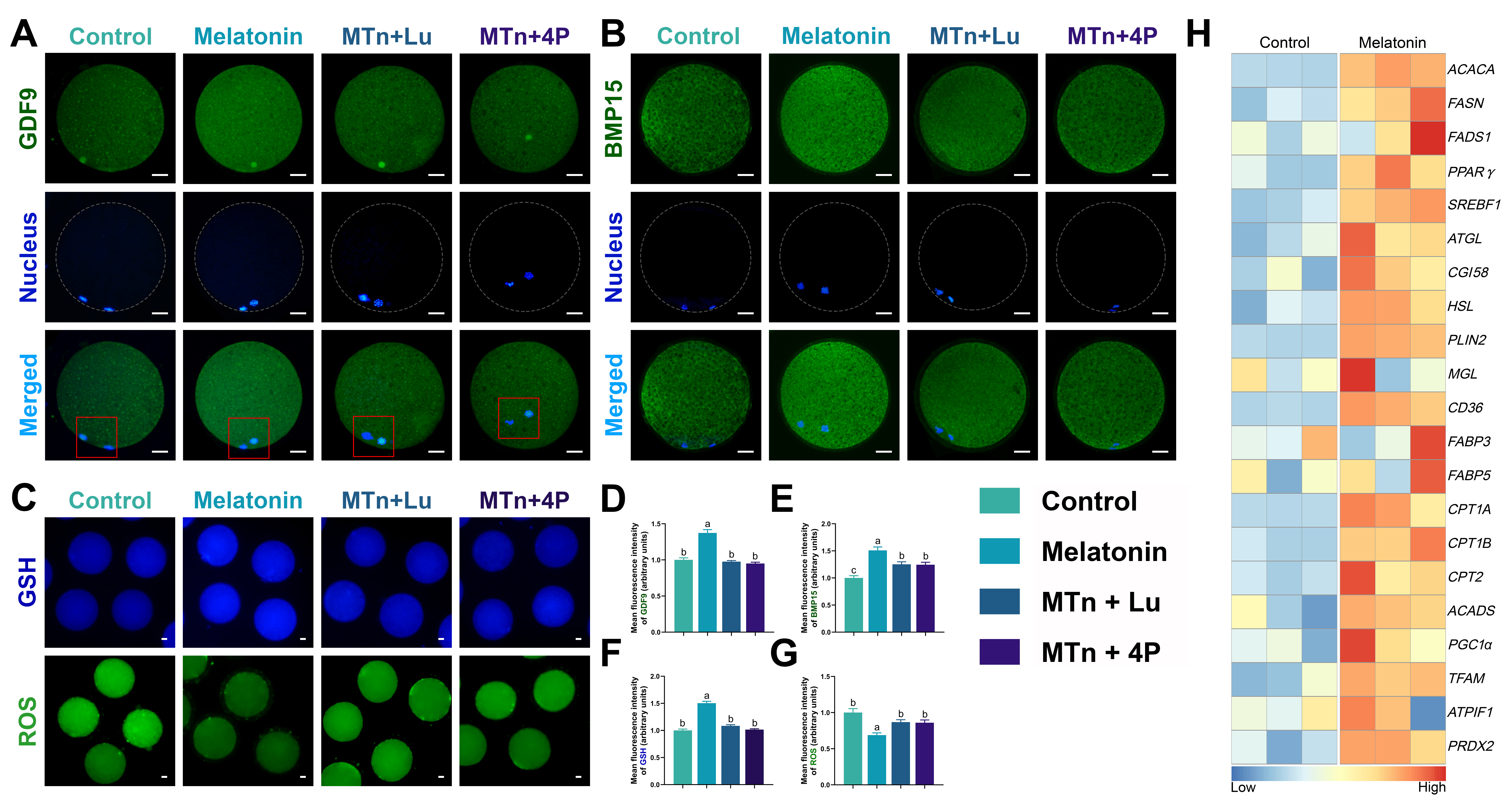
Oocyte maturation is important step for the ovum to reach competence for successful insemination and early embryonic development, and is mainly regulated by bilateral interactions of cumulus cells and oocyte [11, 30]. The oocytes without cumulus cells exert negatively influences to oocyte nuclear maturation, cytoplasmic maturation and early embryonic development after fertilization [31]. Hence, cumulus cells are critical in supporting oocyte maturation during IVM. In the previous study, we have observed the melatonin receptors (MT1 and MT2) expression in porcine cumulus cells [32]. In the present study, we found that protein of MT1 and MT2 could be detected in cumulus cells by IF and WB, and that their expressions were significantly enhanced by melatonin supplementation. Additionally, the melatonin-mediated increase in MT2 expression was blocked by luzindole or 4P-PDOT, while melatonin-mediated MT1 expression was completely eliminated by luzindole, but only partially by 4P-PDOT. Indeed, 4P-PDOT plays its own specific role in COC development, so these results supported our subsequent experiments to research the function of MT1 and/or MT2 in cumulus cells.
Adenylate cyclase (AC) is primarily activated downstream of GPCRs by interaction with the α subunit of the Gs protein [33]. The second messenger (cAMP) was generated by AC activation, it could regulate several effectors, the most well studied of which is cyclic adenosine monophosphate (cAMP)-dependent protein kinase A (PKA) [34]. The enzyme of PKA is participated numerous pathological and physiological actions in mammals and control homeostasis in eukaryotes [35, 36]. It has been indicated that melatonin receptors (MT1 and/or MT2) could regulate intracellular processes via AC, which accumulates cAMP concentration, stimulating PKA activity in human granulosa-lutein cells [37]. In addition, HSL is translocated from the ooplasm to the LD surface following PKA stimulation, where it interacts with the PLIN family in stimulated lipolysis [38, 39]. After activation of PKA, ATGL also interacts with PLIN, despite this most probably takes place indirectly through the co-factors CGI58 in adipocytes [40]. The lipolysis of LDs in COCs is also catalyzed by lipolytic enzyme including HSL and ATGL [41]. Moreover, HSL also catalyzes the lipolysis of diglycerides to monoglycerides, suggesting that it has a stronger lipolytic function than ATGL [42–44]. In the current study, melatonin significantly increased Gsα expression, which up-regulated the cAMP/PKA pathway, and then activated the lipolytic processes with promoting lipases (ATGL and HSL) and their co-factors (PLIN and CGI58) in cumulus cells. Consequently, melatonin significantly decreased the number of intracellular LDs, and increased FA and ATP levels, and enhanced genes transcripts related to β-oxidation in cumulus cells. Nevertheless, these melatonin-mediated effects were completely blocked by luzindole or 4P-PDOT. Based on these results, it appeared that the melatonin-mediated Gsα-PKA signaling pathway could facilitate the lipolytic processes via MT2, producing metabolic substrates as essential energy for cumulus cell development during IVM. In our previous study, we elucidated that the cumulus cells expansion and embryonic development were increased by melatonin treatment via the sonic hedgehog signaling activation [32]. It is extremely likely that the sonic hedgehog signaling was regulated by MT2, because the activation of PKA leads to phosphorylation and stimulation of Smoothened (Smo) [45]. He et al. have demonstrated that the function of porcine granulosa cells was modulated by MT2 receptor [46], and it is suggested that MT2 is likely to mediate porcine cumulus cell expansion by melatonin effects.
As reported previously, a porcine oocyte contains 161 ± 18 ng of endogenous lipid content, the majority of which is in the TG form, much more than for other species [13, 47]. Therefore, porcine oocytes are widely recognized as an excellent model to study the role of lipid metabolism in mammalian female germ cells. Generally, the intracellular lipids in the oocyte are mainly stored in LDs and released FAs by activation of lipolysis, which provide energy for oocyte maturation and development [9, 13, 48, 49]. Additionally, it is recognized that FAs have powerful anti-inflammatory property [50] such as specialized proresolving lipid mediators (resolvins and protectins) which combined with specific G-protein-coupled receptors and reduced hydrogen peroxide-induced DNA damage and oxidative stress [51–53]. We have demonstrated previously that melatonin enhanced lipogenesis- and lipolysis-related mRNA expression in the porcine oocyte [9]. Continuing from these results, we detected several lipogenetic and lipolytic proteins in oocytes, the levels of which were significantly increased by melatonin treatment, leading to increased FA and ATP generation. Thus, melatonin-mediated lipid metabolism dramatically improved oocyte quality and blastocyst formation rate by decreasing ROS and increasing GSH levels, though not nuclear maturation. Further, mRNA expression of CD36, but not FABP3 and FABP5, was significantly increased by melatonin treatment. Thus, the FA transportation-mediated lipid accumulation was mediated by the CD36 and lipogenesis process; and that this was the energy source generated to support oocyte and embryo development. Moreover, the removal of cumulus cells during IVM induced an imbalance in lipid metabolism, affecting the developmental competence of the oocyte [54, 55]. Therefore, these results indicated that the cumulus cells not only protected the developing oocyte but also maintained lipid homeostasis by supporting intracellular lipogenetic and lipolytic processes in the oocyte.
{kind=link}
{kind=link}