Chronic wasting disease (CWD) is a transmissible spongiform encephalopathy caused by prions that has spread across cervid species in North America since the 1960s and recently spread to cervids in Eurasia. The Association of Zoos and Aquariums (AZA) considers CWD to be of major concern for cervids in AZA-accredited facilities because of the indirect transmission risk of the disease and the impact of CWD regulatory protocols on captive breeding programs. Vulnerability to CWD is affected by variation in the PRNP gene that encodes the prion protein. We therefore sequenced PRNP in Pere David’s deer (Elaphurus davidianus), a species that was extinct in the wild for more than a century, and descends from ca. 11 founders. In 27 individuals, we detected two PRNP haplotypes, designated Elad1 (51 of 54 chromosomes) and Elad2 (3 of 54 chromosomes). The two haplotypes are separated by four single nucleotide polymorphisms (SNPs), three of which are non-synonymous. Both Elad1 and Elad2 have polymorphisms that in other cervid taxa are associated with reduced vulnerability to CWD. The two haplotypes are more similar in sequence to PRNP in other cervids than to each other. This suggests that PRNP in cervids may have been under long-term balancing selection, as has been shown for PRNP in non-cervid taxa, and which could account for the presence of multiple haplotypes among founders. There may be a fitness benefit in maintaining both PRNP haplotypes in the species because variation in the prion amino acid sequence can limit transmission of CWD.
Research Article
Variation In The PRNP Gene Of Pere David’s Deer (Elaphurus Davidianus) May Impact Genetic Vulnerability To Chronic Wasting Disease
https://doi.org/10.21203/rs.3.rs-566057/v1
This work is licensed under a CC BY 4.0 License
Journal Publication
published 24 Nov, 2021
You are reading this latest preprint version
cervids
encephalopathy
milu
transmissible spongiform encephalopathy
Chronic wasting disease (CWD) is a highly transmissible spongiform encephalopathy (TSE) caused by prions (Belay et al. 2004; Williams and Young 1980). Prions are “proteinaceous infectious particles” that arise when a normal cellular prion protein (PrPC) misfolds into a protein with an abnormal yet stable protein structure (Belay et al. 2004; Prusiner SB 1991; Prusiner S. B. 1982). This abnormal structure (PrPCWD) causes the cellular prion protein to become infectious, binding to a normal PrPC to induce conformational changes to render it an abnormal PrPCWD. This transformation destroys the cellular protein's standard function in the central and peripheral nervous systems and renders it inactive (Wulf et al. 2017). Prions build up over time, which causes the deterioration of the organism’s nervous system due to the accumulation of the insoluble PrPCWD aggregates. These insoluble aggregates resist degradation by proteases, and are toxic, leading to cell death, and the sponge-like damage to brain tissue that characterizes transmissible spongiform encephalopathies (TSEs). CWD infection is invariably fatal, with no known treatment or cure (Belay et al. 2004; Williams and Young 1980). CWD can be spread via direct animal to animal contact (infectious to susceptible animal) through exchange of bodily fluids such as saliva, urine, blood, and semen (Haley et al. 2011; Haley et al. 2009; Kramm et al. 2020; Mathiason et al. 2006), although direct contact with wild cervids is a less likely occurrence for deer kept in most zoos than for deer kept in deer ranches. Prions are also sequestered by plants and remain infectious in certain types of soil particles for years (Bartelt-Hunt and Bartz 2013; Kuznetsova et al. 2018; Kuznetsova et al. 2014; Nichols et al. 2009). They can also remain infectious on a variety of materials, including glass, plastics, and steel instruments even after autoclaving (Edgeworth et al. 2009; Pritzkow et al. 2018). Environmental CWD transmission to mule deer from paddocks that previously held infected animals has been experimentally confirmed as well (Miller et al. 2004). Epidemiological models indicate that the indirect transmission risk of contracting CWD is substantial (Almberg et al. 2011; Saunders et al. 2012).
CWD was initially characterized in the 1960s in Colorado and has since spread geographically in North America across populations of both free-ranging and captive cervids in at least 24 U.S. states and two Canadian provinces (CDC 2020; Richards 2020; Rivera et al. 2019; Williams and Young 1980; Williams and Young 1982). U.S. federal and state agencies (including the U.S. Fish and Wildlife Service) actively manage, monitor, and are attempting to reduce the spread of CWD (Manjerovic et al. 2014; Mateus-Pinilla et al. 2013). Despite these efforts, CWD has recently spread to wild cervid populations in Finland, Norway and Sweden, and captive populations in South Korea (Richards 2020). Global concern for CWD containment has increased because CWD may cause population declines after becoming endemic in a cervid population (Edmunds et al. 2016).
The increasing geographic spread of CWD has raised worries about its potential to impact the health of endangered cervids in the wild or captive breeding programs (CDC 2020). Small populations such as those kept in captive breeding programs are particularly vulnerable to the impact of infectious disease (Cunningham et al. 2017; Viggers et al. 1993). The Association of Zoos and Aquariums (AZA), has established guidelines for the monitoring of CWD, and for the management of cervids that could be susceptible to CWD. These guidelines include restrictions on the transfer of cervids from non-accredited facilities which may not be monitoring for CWD in their cervid populations, strict protocols regarding the interstate transport of cervids, necropsy surveillance of deceased cervids in facilities, and measures to reduce risk from free-ranging cervids (AZA Board of Directors 2003). However, CWD can and has been unintentionally transmitted to captive populations via infected feed and bedding and not just from direct transmission by infected animals (Henderson et al. 2015; Mathiason et al. 2009; Mathiason et al. 2006) indicating that cervids in AZA-accredited institutions are at risk. CWD has also been detected in cervid semen, raising concerns on how this may impact the use of artificial insemination in captive cervid facilities (Kramm et al. 2020).
Another interest of AZA-accredited institutions is to better understand the risk of CWD transmission to determine if current protocols regarding the transport of cervids are appropriate. Strict restrictions on interstate transfers, such as those in place in many states, can hinder cervid breeding programs in AZA-accredited facilities. Thus, further elucidating transmission risk is important so that more precise protocols and restrictions can be established. The AZA Cervid Taxon Advisory Group monitors cervid species in captive breeding programs, including the Pere David’s deer (Elaphurus davidianus), which has been categorized by the IUCN red list (https://www.iucnredlist.org) as extinct in the wild.
The deer family Cervidae is divided into the subfamilies Cervinae and Capreolinae (Heckeberg 2020). The subfamily Capreolinae includes the cervid species of North and South America, and some species found in Eurasia (Fernández and Vrba 2005). The subfamily Capreolinae includes white-tailed deer (Odocoileus virginianus), mule deer (Odocoileus hemionus), moose (Alces alces), caribou/reindeer (Rangifer tarandus), roe deer (Capreolus capreolus), and the Chinese water deer (Hydropotes inermis) among others. Pere David’s deer is within the subfamily Cervinae, which also includes the red deer (Cervus elaphas), elk or wapiti (Cervus canadensis), sika deer (Cervus nippon), Eld’s deer (Panolia eldii), fallow deer (Dama dama), spotted deer (Axis axis), and barasingha (Rucervus duvaucelii), among others. This subfamily originated in Eurasia and is characterized by elaborate antlers at obtuse angles and shared plesiometacarpal traits (Pitra et al. 2004).
Pere David’s deer (also called the milu) historically ranged across the swamplands of eastern Asia with the largest populations found in southern mainland China. There are accounts of the species being kept in captivity dating back over a thousand years (Schafer 1968). Pere David’s deer became extinct in the wild in the late 19th century due to overhunting (Schafer 1968; Yuan et al. 2019; Zhu et al. 2018). The species is named after Father Armand David, a French missionary in China who sent specimens to European museums. The last known population within China was kept in the Nanyuang Royal Hunting Garden; these deer were killed during the Boxer Rebellion of 1899-1901 (Jiang et al. 2000; Li et al. 2011; Schafer 1968). The modern population is descended from a herd of 18 deer housed in the United Kingdom at Woburn Abbey, of which 11 are believed to have left descendants (Jones 1951; Zeng et al. 2007; Zhang Chenzhou et al. 2017).
In the 1980s, Pere David’s deer started to be reintroduced as free-ranging herds in China, with the largest reintroduced population (over 3,000 individuals) now residing in the Dafeng coastal region (Hu and Jiang 2002; Jiang et al. 2000; Zhu et al. 2018). However, much of the population still resides in captive facilities, which are used to supplement the free-ranging populations in China (Zhu et al. 2018). Despite being descended from only 11 individuals, Pere David's deer have not exhibited obvious adverse effects indicative of inbreeding depression (Yuan et al. 2019). In 2018, the genomes of five unrelated Pere David’s deer were analyzed. Their genomes had limited runs of homozygosity, and one analysis suggested that the effective population size had declined continuously, which potentially may have allowed the species to be largely purged of deleterious alleles (Zhu et al. 2018). Despite the low number of founders, Pere David’s deer do not seem to exhibit negative genetic effects or the extreme lack of genetic diversity expected of a population that undergoes a severe bottleneck (Zhang Xuejing et al. 2016; Zhu et al. 2018). Pere David’s deer are often mentioned as a model of successful captive population management since the population has rebounded from so few founders (Jiang et al. 2000; Zhu et al. 2018). It has been suggested that having a low juvenile mortality rate and an overall lack of inbreeding depression may have played a large role in Pere David’s deer recovery (Zhu et al. 2018).
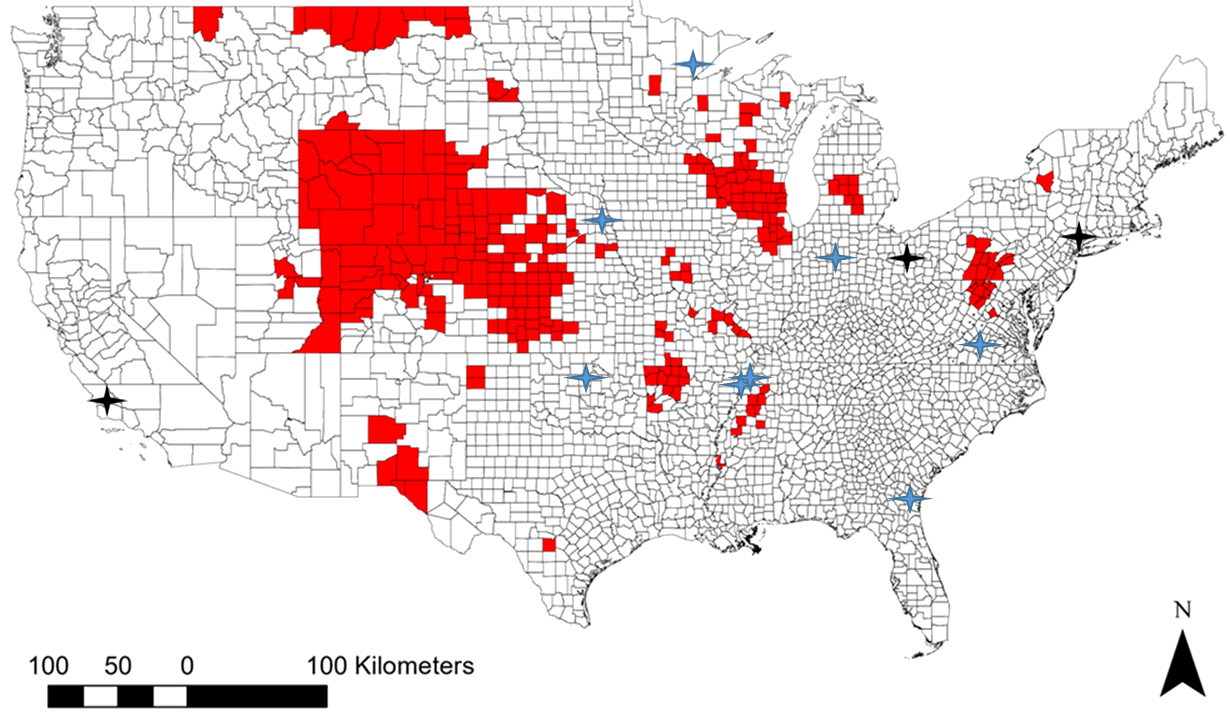
As with other cervids, there is concern that Pere David's deer could be susceptible to CWD. CWD has not yet been reported in counties where AZA-accredited institutions maintain Pere David’s deer in captive breeding programs (Supplementary Figure 1). However, cervids in these facilities are still at risk because CWD can be transmitted through feed and bedding, and direct transmission from cervids brought in from non-accredited captive settings or potential infection from semen during artificial insemination (Henderson et al. 2015; Kramm et al. 2020; Mathiason et al. 2009)
The gene PRNP encodes the prion protein. Genetic variation in PRNP has been associated with substantial differences in vulnerability to CWD in many cervid taxa. For example, there is evidence from fallow deer that asparagine at codon 138 is associated with reduced vulnerability to CWD. Asparagine at codon 138 is associated with reduced CWD vulnerability in caribou as well (Cheng et al. 2017; Haley et al. 2017; Mitchell et al. 2012; Moore et al. 2016; Rhyan et al. 2011). Variation at codons in other taxa has also been associated with differences in CWD vulnerability. While PRNP has been sequenced in two Pere David’s individuals (KC476497 and PRJNA256236), the degree and type of variation has not been broadly examined. Therefore, we here sequenced the entire coding region of PRNP in 27 Pere David’s deer, to inform the conservation and management of the species by examining PRNP variation. Despite the limited number of founders, we detected PRNP variation in Pere David’s deer, including non-synonymous substitutions that in other deer species are associated with reduced susceptibility to CWD.
Pere David’s deer sampling
A total of 27 samples from different Pere David’s deer individuals were provided by AZA-accredited institutions and sequenced for this study. The Wilds, a non-profit safari park and conservation center, located in Cumberland, Ohio, provided 15 samples; the San Diego Zoo Institute for Conservation Research provided six samples; and the Wildlife Conservation Society provided six samples from the Bronx Zoo (Table 1). The samples were from previously stored blood collections or were collected during routine veterinary procedures.
DNA amplification and sequence analysis
DNA was extracted from blood samples using the QIAamp DNA Blood Mini Kit (Qiagen, Germantown, MD) using a one-hour lysis time. Manufacturer’s instructions were followed in all other aspects of the protocol. The DNA was amplified by PCR in 25 μl total volume, containing 1× PCR Buffer II (Applied Biosystems Inc. [ABI]), final concentrations of 200 μM of each of the dNTPs, 1.5 mM MgCl2, 0.04 units/μl of AmpliTaq Gold DNA Polymerase (ABI) and 0.4 μM of each oligonucleotide primer. The forward primer 223 (5’-acaccctctttattttgcag-3’) and the reverse primer 224 (5’-agaagataatgaaaacaggaag-3’) were used to amplify and sequence an 830 bp region encompassing the full open reading frame, encoded by exon 3 of PRNP (O'Rourke et al. 2004). Primers 223 and 224 had been designed to avoid amplifying the PRNP pseudogene that lacks introns (O'Rourke et al. 2004). PCR conditions for PRNP amplification were as follows: 95˚C for 10 minutes for the initial denaturing; 5 cycles of 95˚C for 30 seconds, 56˚C for 30 seconds, and 72˚C for 1 minute; and 40 cycles of 95˚C for 30 seconds, 50˚C for 30 seconds, and 72˚C for 1 minute; with a final extension of 7 minutes at 72˚C. PCR amplification was confirmed with a 1.0% agarose gel using gel electrophoresis. The successful amplification products were enzyme-purified (Hanke and Wink 1994). Sequencing was conducted in both directions, relying on each of the PCR primers, with primers PRNP-IF 5’- atgctgggaagtgccatga-3’ and PRNP-IR 5’-catggcattcccagcat-3’ used as internal primers (Ishida et al. 2020). Sequences were generated using the BigDye Terminator v3.1 Cycle Sequencing Kit (ABI), with 1.0 μl of purified PCR product and 0.12 μM of primer, resolved on an ABI 3730XL DNA Sequencer at the Keck Center for Functional and Comparative Genomics at the University of Illinois at Urbana-Champaign. Sequences were then visually confirmed and concatenated using Sequencher software 5.4.6 (Gene Codes Corporation, Ann Arbor, MI).
Haplotype analysis
Phase v.2 within the software DnaSP was used to phase the sequence data and infer haplotypes (Librado and Rozas 2009; Stephens et al. 2001); 10,000 iterations were run with 1000 burn-in iterations. Haplotype sequences were aligned using Sequencher, which was also used to confirm open reading frames. Haplotype identity was verified using NCBI Blast (https://blast.ncbi.nlm.nih.gov/Blast.cgi). Confidence intervals for the haplotypes were calculated (Hazra 2017) using the following equation: . Sequences were translated using MEGA X v.10.1 (Kumar et al. 2018). The software PopART (with default parameters) was used to generate and illustrate a median-joining network (Bandelt et al. 1999; Leigh 2015). The haplotype sequences generated by this study were deposited in GenBank (accession numbers MW804582 and MW804583).
. Sequences were translated using MEGA X v.10.1 (Kumar et al. 2018). The software PopART (with default parameters) was used to generate and illustrate a median-joining network (Bandelt et al. 1999; Leigh 2015). The haplotype sequences generated by this study were deposited in GenBank (accession numbers MW804582 and MW804583).
For all 27 Pere David’s deer available, DNA was successfully extracted, and the complete open reading frame of PRNP was amplified and sequenced (Table 1). Four single nucleotide polymorphisms (SNPs) and two haplotypes were identified in the coding region. The two haplotypes detected were designated Elad1 and Elad2. The DNA sequence of haplotype Elad1 is the same as two previously reported Pere David’s deer PRNP sequences (GenBank accession numbers: KC476497 and PRJNA256236). The Pere David’s deer PRNP haplotype Elad2 sequence is novel and has not been previously detected in any other cervid species, nor has its PrP amino acid sequence been previously reported.
Haplotype Elad1 was detected in 51 of 54 (0.944 ± 0.061 [95% confidence interval or CI]) phased Pere David’s deer sequences (i.e., chromosomes), and thus had the highest frequency. Haplotype Elad2 had a much lower frequency and was carried by three of the 54 Pere David’s deer chromosomes examined (95% CI: 0.056 ± 0.0612). Samples of deer carrying Elad2 were extracted, amplified and sequenced a second time for verification. Haplotypes Elad1 and Elad2 were separated by four SNPs. One of the polymorphisms, c.183C>T is a synonymous SNP which has not been previously detected across cervids. The three other SNPs were non-synonymous SNPs encoding amino acid variation previously detected in other species of cervids (Table 2). In Elad2, SNP c.413A>G encodes serine (S) at codon 138, which is also present in many deer taxa (Table 2); whereas in Elad2, asparagine (N) is encoded at codon 138 (as reported in fallow deer and some caribou; Table 2) (Cheng et al. 2017; Haley et al. 2017; Moore et al. 2016; Rhyan et al. 2011). SNP c.624G>A encodes isoleucine (I) at codon 208 in Elad2, whereas methionine (M) is encoded by Elad1. Both variants have been reported in red deer (Cervus elaphus) (Kaluz et al. 1997), whereas methionine is common in other deer species (Table 2). SNP c.676C>G encodes glutamine (Q) at codon 226 in Elad2, whereas glutamic acid (E) is encoded by Elad1; both variants have been reported in red deer and sika deer (Table 2) (Haley et al. 2017; Jeong et al. 2007; Robinson et al. 2019).
Amino acid sequences of PrP were aligned for some cervid taxa with PRNP haplotype sequence available in GenBank, along with Elad1 and Elad2 (Table 2). These sequences represent much of the reported PrP variation in cervids, and include PrP amino acid variants associated with reduced vulnerability to CWD in cervid taxa, as indicated in Table 2. Both of the Pere David’s haplotypes encoded amino acids that had previously been associated with reduced vulnerability to CWD in other species.
For the cervid taxa shown in Table 2, a median-joining network (Figure 1) was generated using PRNP sequences. The two Pere David’s deer PRNP haplotypes were not most similar to each other. Instead, Elad1 was most similar to a PRNP sequence for fallow deer, while Elad2 was more similar to a PRNP sequence that is shared by Iberian red deer (Cervus elaphus hispanicus), Rocky Mountain elk (Cervus elaphus nelsoni), and sika deer carrying the 226E substitution.
No CWD infections have so far been reported among Pere David’s deer. Although direct transmission from free-ranging deer may be possible at some facilities, the most likely potential means of transmission of CWD to Pere David’s deer stocks in North America would be through feed or bedding. These can be contaminated by CWD-positive free-ranging North-American cervids, which can shed prions that persist in the environment and remain infectious (Henderson et al. 2015; Mathiason et al. 2009). There is also the concern that infected semen can be an issue for captive facilities that use artificial insemination on their cervid populations, although the transmission risk is not currently known (Kramm et al. 2020).
Given the risk of transmission of CWD, it is worth comparing the amino acids sequence of Pere David’s deer PrP with those of North American cervids undergoing outbreaks of CWD (Table 2, Figure 1). In white-tailed deer, the SNP c.285A>C, which encodes a histidine (H) at codon 95, and the SNP c.286G>A, which encodes a serine (S) at codon 96, are associated with reduced susceptibility to CWD in white-tailed deer (Brandt et al. 2018; Brandt et al. 2015; Ishida et al. 2020; Johnson et al. 2003; Kelly et al. 2008; O'Rourke et al. 2004). Mule deer with at least one chromosome in which PRNP encodes phenylalanine (F) at codon 225 show a reduced frequency CWD (Geremia et al. 2015; Jewell et al. 2005). Pere David’s deer do not encode any of these three amino acids that are associated with reduced susceptibility to CWD in the native North American cervid species. However, this is not an indication of high vulnerability to CWD among Pere David’s deer, as other codons present in the Pere David’s deer sequence differed from those present in North American species.
The Elad1 and Elad2 haplotypes carried by Pere David’s deer are separated by three non-synonymous SNPs in addition to a synonymous SNP. Elad1, the haplotype present in 51 of the 54 chromosomes, encoded asparagine (N) at codon 138, methionine (M) at codon 208 and glutamic acid (E) at codon 226; whereas haplotype Elad2, present in three of 54 chromosomes, encoded serine (S), isoleucine (I), and glutamine (Q), respectively (Table 2). All substitutions are conservative replacements (Betts 2003).
Both serine (S) and asparagine (N) at codon 138 are neutrally charged polar amino acids, but serine is smaller than asparagine, suggesting that this substitution could potentially cause substantial configurational changes (Betts 2003). Elad1 encodes asparagine (N) at codon 138. This polymorphism has been associated with lower susceptibility to CWD in fallow deer and caribou (Cheng et al. 2017; Haley et al. 2017; Mitchell et al. 2012; Moore et al. 2016; Rhyan et al. 2011), species in different subfamilies of the Cervidae (Table 2). In caribou, several studies have indicated that deer that carry at least one chromosome that encodes N at codon 138 show reduced vulnerability to CWD or slower infection rates. The S138N substitution confers reduced susceptibility in experimental inoculation studies, and there was significantly lower CWD in caribou herds with high 138N frequencies (Cheng et al. 2017; Happ et al. 2007; Mitchell et al. 2012). In fallow deer (n=66), the S138N substitution appears to be fixed (Robinson et al. 2019). In-vitro conversion of cervid PrP from elk, white-tailed deer, and mule deer (PrP variants from each of these taxa were cloned and expressed in cultured cells) into an infectious PrPCWD form is reported to be more efficient for 138S than for the 138N (Raymond et al. 2000). This finding suggests that the S138N substitution provides reduced vulnerability to CWD (Raymond et al. 2000). Elad1 encodes 138N, which suggests that it would also have a protective role in Pere David’s deer. However, experiments would have to be directly conducted on the species itself (in vivo or in vitro) to establish whether the substitution may provide protection among Pere David’s deer.
Both methionine (M) and isoleucine (I) at codon 208 are relatively unreactive and hydrophobic (Betts 2003). The substitution of methionine for isoleucine has been reported not to cause substantial structural changes to proteins (Ohmura et al. 2001). Despite this being a conservative substitution, isoleucine at codon position 208 appears to play a significant role in preventing in vitro interspecies transmission of CWD from deer to sheep (sheep encode isoleucine at codon 208; while the cervid PrP in the Harrathi study had methionine) (Harrathi et al. 2019). Elad1 carries 208M as do the listed North American deer species, while Elad2 encodes isoleucine at codon 208 (Table 2). Thus, it is possible (though not established) that 208I present in Elad2 may reduce vulnerability to CWD of Pere David’s deer carrying this haplotype if exposed to CWD from North American deer that do not carry this substitution. There is also the potential for reduced transmission between Pere David’s deer carrying Elad1 and those carrying Elad2 due to potential barriers in transmission of CWD to PrP of variant configurations (Kurt and Sigurdson 2016). Both of these possibilities would suggest that the two haplotypes should be maintained in Pere David’s deer populations, pending additional species-specific studies.
The final non-synonymous substitution within Pere David’s deer is glutamine (Q) to glutamic acid (E) at codon 226. This is a common amino acid substitution, as both are polar and have similar physiochemical properties (Betts 2003). There is variation in the reported effects of the Q226E polymorphism in different cervid taxa. In red deer, deer homozygous for both the 226E and 226Q substitutions as well as heterozygous deer contracted CWD from an oral inoculation experiment with no significant difference in vulnerability (Balachandran et al. 2010). A study conducted in transgenic mice compared mice expressing a PrP variant found in Rocky Mountain elk with mice expressing a PrP variant found in both white-tailed deer and mule deer. It found that that 226E carried by Rocky Mountain elk might allow for more efficient CWD prion propagation than the 226Q carried by mule deer and white-tailed deer (Angers et al. 2010). Yet, all reported fallow deer carry the 226E substitution (along with the 138N substitution), and are still resistant to direct infection by CWD from other cervids or indirect infection by a contaminated environment (Rhyan et al. 2011). As the role of the Q226E polymorphism on CWD vulnerability varies by species, studies for individual species are needed to determine what role if any it may play in vulnerability to CWD for each taxon. Pere David’s deer Elad2 encodes 226E, which in native North American cervids has been reported only among Rocky Mountain elk; other North American taxa carry 226Q, which is also present in Elad1 (Table 2).
These previous findings suggest that if Pere David’s deer populations were to be exposed to the CWD present in native North American cervids, there may be reduced vulnerability due to differences in proteins encoded both by Elad1 and Elad2. Elad1 carries 138N, which reduces CWD vulnerability in both fallow deer and caribou (Cheng et al. 2017; Haley et al. 2017; Mitchell et al. 2012; Moore et al. 2016; Rhyan et al. 2011). Elad2 carries 226E, which has been associated with relatively higher CWD vulnerability in Rocky mountain elk and no effect on vulnerability in other species. However, Elad2 also encodes 208I, which might provide reduced vulnerability to CWD (Angers et al. 2010; Balachandran et al. 2010; Harrathi et al. 2019; Rhyan et al. 2011). Again, it must be emphasized that the studies establishing a role for these three non-synonymous substitutions were conducted on other species, and a similar reduction in vulnerability to CWD would have to be established by in vitro or in vivo studies of Pere David’s deer. Thus, all guidelines to reduce the potential spread of CWD in captive breeding programs for cervids should continue to be followed for Pere David’s deer (Cullingham et al. 2020).
Despite the small number of founders, the two distinctive haplotypes have persisted into the modern population of Pere David’s deer (Figure 1). Both Elad1 and Elad2 share PrP amino acids with related cervid species (Table 2). The presence of trans-species polymorphisms, i.e., similar substitutions shared by different species, can be indicative of long-term balancing selection. The alleles that distinguish Elad1 from Elad2 were both likely present in an ancestor that gave rise to different species of the subfamily Cervinae (Charlesworth 2006; Klein et al. 1998; Koenig et al. 2019), and have since persisted in Pere David’s deer. Balancing selection driven by transmissible spongiform encephalopathies has been identified for PRNP in other species, including human populations exposed to kuru (Mead et al. 2003), and sheep (Ovis aries) populations exposed to scrapie (Slate 2005). In North American elk, populations exposed to CWD may be under balancing selection at the codon 132, which extends CWD latency but also potentially has negative fitness consequences (Monello et al. 2017). The presence of ancestral trans-species polymorphisms, and of multiple haplotypes in the small founder population, would both be consistent with the ancestors of Pere David’s deer being subject to balancing selection at the PRNP gene. If the persistence of Elad1 and Elad2 in Pere David’s deer is due to balancing selection, this would suggest a fitness benefit for maintaining both haplotypes in the population. The frequency of Elad2 should be monitored so that it persists in Pere David’s deer stocks. However, given the presence of 138N in Elad1 and the strong evidence for its protective role in fallow deer and caribou, it should be maintained as the more common haplotype.
The maintenance of both haplotypes in the Pere David’s deer population is also recommended due to a likely reduction in potential intra-species transmission between deer carrying different PrP proteins, should CWD enter the population. Variation in CWD strain and PrP primary configuration have been shown to change PrP conversion rates and thus the presence of both PrP variants within a population could alter the effectiveness or speed of a CWD infection (Collinge and Clarke 2007; Kurt and Sigurdson 2016). Genetic management of captive populations of Pere David’s deer to maintain both haplotypes should be considered an additional strategy for CWD management. This could be undertaken without affecting other goals, such as equalizing founder contributions; for example, if only one of two full siblings (which have identical ancestors) carries Elad2, then it could be favored over the other sibling for reproduction. Gametes from males that carry the Elad2 haplotype could also be collected and stored, to be used for the breeding of Pere David’s deer in the future. The rarity of Elad2 suggests that increasing its frequency may have favorable fitness consequences if either heterozygote advantage or rare allele effects are relevant to PRNP. Given the potentially protective role of 138N in Elad1, deer could also be selectively bred to maintain Elad2 in a heterozygous state.
We present an assessment of PRNP genetic polymorphisms in Pere David’s deer, identifying two haplotypes including the novel Elad2. The large majority (94.4%) of Pere David’s deer chromosomes carried haplotype Elad1, which encodes asparagine (N) at codon 138 and glutamine (Q) at codon 226, both associated with a potential reduction in CWD susceptibility based on studies in other species. The remaining chromosomes carried Elad2, with isoleucine at codon 208, associated with lower susceptibility to CWD in another cervid species. Thus, it is possible that Pere David’s deer may have reduced vulnerability to CWD. The trans-species polymorphisms detected across cervid species, and the presence of two divergent haplotypes among the small number of Pere David’s deer founders are both potential signatures of ancestral balancing selection. Pere David’s deer should be managed through breeding programs to prevent the loss of rare haplotype Elad2, since the presence of two distinctive variants of PrP may help to limit the spread of CWD among the species, should there be an outbreak.
It seems unlikely, given the small number of founders, that additional PRNP haplotypes would be detected in Pere David’s deer. However, additional individuals should be sequenced to identify those with Elad2. Wildlife ranches that have free-ranging herds are being considered as a source of breeding stock to add to the genetic diversity and conservation aims of zoos (Wildt et al. 2019). These free-ranging deer could be assessed for the presence of Elad2 (or additional haplotypes). The sequencing of PRNP in Pere David’s deer has helped to characterize the potential genetic vulnerability of this species to CWD. Similar surveys of PRNP genetic variation on other endangered cervid taxa may be conducted to assess their genetic vulnerability to CWD. Finally, inferences of the risk posed by CWD to Pere David’s deer were based on studies conducted on other species. Additional studies in vivo or in vitro studies involving Pere David’s deer, to directly examine the risk of various CWD strains to the species, would provide more direct information on the risk to Pere David’s deer. Such studies could also help quantify the relative risk for homozygotes of Elad1 and Elad2, and for heterozygotes, which in turn could be used to determine ideal frequencies for the two haplotypes in the captive breeding stock of Pere David’s deer.
Funding: This work was supported by the Cooperative State Research, Education, and Extension Service, U.S. Department of Agriculture, under project number ILLU 875-952 and ILLU 538-966. This work was also supported by the Francis M. and Harlie M. Clark Research Support Grant.
Conflicts of interest: The authors declare no conflict of interest in presenting this publication.
Availability of data and material: PRNP haplotype sequences for Pere David’s deer have been deposited in GenBank under accession numbers MW804582 and MW804583. Sample information is listed in Table 1.
Code availability: Not applicable
Ethics approval: Research was conducted under the Illinois Institutional Animal Care and Use Committee protocol 18212. Individual material transfer agreements are in place for each institution that contributed samples, and all terms were adhered to.
Consent to participate (include appropriate statements): Not applicable
Consent for publication (include appropriate statements): Not applicable
Acknowledgments
For providing Pere David’s deer samples, we thank The Wilds in Cumberland, Ohio; the San Diego Zoo Institute for Conservation Research in San Diego, California; and the Wildlife Conservation Society and the Bronx Zoo in Bronx, New York. We would like to thank Dr. Dee McAloose, Steve Metzler, Dr. Jean Paré, Dr. Cynthia Steiner and anyone else who was involved in providing sample information and assisting in sample preparation and collection. We thank Alida de Flamingh for assistance in designing supplementary figure 1.
- Almberg ES, Cross PC, Johnson CJ, Heisey DM, Richards BJ (2011) Modeling routes of chronic wasting disease transmission: environmental prion persistence promotes deer population decline and extinction. PLoS One 6:e19896. https://doi.org/10.1371/journal.pone.0019896
- Angers R, Kang HE, Napier D, Browning S, Seward T, Mathiason C, Balachandran A, McKenzie D, Castilla J, Soto C, Jewell J, Graham C, Hoover EA, Telling GC (2010) Prion strain mutation determined by prion protein conformational compatibility and primary structure. Science 328:1154–1158. https://doi.org/10.1126/science.1187107
- AZA Board of Directors A (2003) Guidelines for chronic wasting disease surveillance. https://www.aza.org/guidelines-for-chronic-wasting-disease-surveillance
- Balachandran A, Harrington NP, Algire J, Soutyrine A, Spraker TR, Jeffrey M, González L, O'Rourke KI (2010) Experimental oral transmission of chronic wasting disease to red deer (Cervus elaphus elaphus): early detection and late stage distribution of protease-resistant prion protein. Can Vet J 51:169–178
- Bandelt HJ, Forster P, Röhl A (1999) Median-joining networks for inferring intraspecific phylogenies. Mol Biol Evol 16:37–48. https://doi.org/10.1093/oxfordjournals.molbev.a026036
- Bartelt-Hunt SL, Bartz JC (2013) Behavior of prions in the environment: implications for prion biology. PLOS Pathogens 9:e1003113. https://doi.org/10.1371/journal.ppat.1003113
- Belay ED, Maddox RA, Williams ES, Miller MW, Gambetti P, Schonberger LB (2004) Chronic wasting disease and potential transmission to humans. Emerg Infect Dis 10:977–984. https://dx.doi.org/10.3201/eid1006.031082
- Betts MJ, Russell RB (2003) Amino acid properties and consequences of substitutions. In: Bioinformatics for Geneticists. John Wiley & Sons, Ltd, https://doi.org/10.1002/9780470059180.ch13
- Brandt AL, Green ML, Ishida Y, Roca AL, Novakofski J, Mateus-Pinilla NE (2018) Influence of the geographic distribution of prion protein gene sequence variation on patterns of chronic wasting disease spread in white-tailed deer (Odocoileus virginianus). Prion 12:204–215. https://doi.org/10.1080/19336896.2018.1474671
- Brandt AL, Kelly AC, Green ML, Shelton P, Novakofski J, Mateus-Pinilla NE (2015) Prion protein gene sequence and chronic wasting disease susceptibility in white-tailed deer (Odocoileus virginianus). Prion 9:449–462. https://doi.org/10.1080/19336896.2015.1115179
- CDC (2020) Chronic wasting disease (CWD): occurrence. Centers for Disease Control and Prevention. https://www.cdc.gov/prions/cwd/occurrence.html
- Charlesworth D (2006) Balancing selection and its effects on sequences in nearby genome regions. PLOS Genetics 2:e64. https://doi.org/10.1371/journal.pgen.0020064
- Cheng YC, Musiani M, Cavedon M, Gilch S (2017) High prevalence of prion protein genotype associated with resistance to chronic wasting disease in one Alberta woodland caribou population. Prion 11:136–142. https://doi.org/10.1080/19336896.2017.1300741
- Collinge J, Clarke AR (2007) A general model of prion strains and their pathogenicity. Science 318:930–936. https://doi.org/10.1126/science.1138718
- Cullingham CI, Peery RM, Dao A, McKenzie DI, Coltman DW (2020) Predicting the spread-risk potential of chronic wasting disease to sympatric ungulate species. Prion 14:56–66. https://doi.org/10.1080/19336896.2020.1720486
- Cunningham AA, Daszak P, Wood JLN (2017) One Health, emerging infectious diseases and wildlife: two decades of progress? Philos Trans R Soc Lond B Biol Sci 372:20160167. https://doi.org/10.1098/rstb.2016.0167
- Doolittle DP (1987) Equilibrium under selection. In: Population Genetics: Basic Principles. Springer Berlin Heidelberg, Berlin, pp 69–73. https://doi.org/10.1007/978-3-642-71734-5_14
- Edgeworth JA, Jackson GS, Clarke AR, Weissmann C, Collinge J (2009) Highly sensitive, quantitative cell-based assay for prions adsorbed to solid surfaces. PNAS 106:3479–3483. https://doi.org/10.1073/pnas.0813342106
- Edmunds DR, Kauffman MJ, Schumaker BA, Lindzey FG, Cook WE, Kreeger TJ, Grogan RG, Cornish TE (2016) Chronic wasting disease drives population decline of white-tailed deer. PLoS One 11:e0161127. https://doi.org/10.1371/journal.pone.0161127
- Fernández MH, Vrba ES (2005) A complete estimate of the phylogenetic relationships in Ruminantia: a dated species-level supertree of the extant ruminants. Biol Rev 80:269–302. https://doi.org/10.1017/S1464793104006670
- Geremia C, Hoeting JA, Wolfe LL, Galloway NL, Antolin MF, Spraker TR, Miller MW, Hobbs NT (2015) Age and repeated biopsy influence antemortem PrP (CWD) testing in mule deer (Odocoileus hemionus) in Colorado, USA. J Wildl Dis 51:801–810. https://doi.org/10.7589/2014-12-284
- Haley NJ, Mathiason CK, Carver S, Zabel M, Telling GC, Hoover EA (2011) Detection of chronic wasting disease prions in salivary, urinary, and intestinal tissues of deer: potential mechanisms of prion shedding and transmission. J Virol 85:6309–6318. https://doi.org/doi:10.1128/JVI.00425-11
- Haley NJ, Rielinger R, Davenport KA, apos, Rourke K, Mitchell G, Richt JA (2017) Estimating chronic wasting disease susceptibility in cervids using real-time quaking-induced conversion. J Gen Virol 98:2882–2892. https://doi.org/10.1099/jgv.0.000952
- Haley NJ, Seelig DM, Zabel MD, Telling GC, Hoover EA (2009) Detection of CWD prions in urine and saliva of deer by transgenic mouse bioassay. PLoS One 4:e4848–e4848. https://doi.org/10.1371/journal.pone.0004848
- Hanke M, Wink M (1994) Direct DNA sequencing of PCR-amplified vector inserts following enzymatic degradation of primer and dNTPs. Biotechniques 17:858–860
- Happ G, Huson H, Beckmen K, Kennedy L (2007) Prion protein genes in caribou from Alaska. J Wildl Dis 43:224–228. https://doi.org/10.7589/0090-3558-43.2.224
- Harrathi C, Fernández-Borges N, Eraña H, Elezgarai SR, Venegas V, Charco JM, Castilla J (2019) Insights into the bidirectional properties of the sheep-deer prion transmission barrier. Mol Neurobiol 56:5287–5303. https://doi.org/10.1007/s12035-018-1443-8
- Hazra A (2017) Using the confidence interval confidently. J Thorac Dis 9:4125–4130. https://doi.org/10.21037/jtd.2017.09.14
- Heckeberg NS (2020) The systematics of the Cervidae: a total evidence approach. PeerJ 8:e8114–e8114. https://doi.org/10.7717/peerj.8114
- Henderson DM, Denkers ND, Hoover CE, Garbino N, Mathiason CK, Hoover EA (2015) Longitudinal detection of prion shedding in saliva and urine by chronic wasting disease-infected deer by real-time quaking-induced conversion. J Virol 89:9338–9347. https://doi.org/10.1128/JVI.01118-15
- Hu H, Jiang Z (2002) Trial release of Père David's deer Elaphurus davidianus in the Dafeng Reserve, China. Oryx 36:196–199. https://doi.org/10.1017/S0030605302000273
- Ishida Y, Tian T, Brandt AL, Kelly AC, Shelton P, Roca AL, Novakofski J, Mateus-Pinilla NE (2020) Association of chronic wasting disease susceptibility with prion protein variation in white-tailed deer (Odocoileus virginianus). Prion 14:214–225. https://doi.org/10.1080/19336896.2020.1805288
- Jeong HJ, Lee JB, Park SY, Song CS, Kim BS, Rho JR, Yoo MH, Jeong BH, Kim YS, Choi IS (2007) Identification of single-nucleotide polymorphisms of the prion protein gene in sika deer (Cervus nippon laiouanus). J Vet Sci 8:299–301. https://doi.org/10.4142/jvs.2007.8.3.299
- Jewell JE, Conner MM, Wolfe LL, Miller MW, Williams ES (2005) Low frequency of PrP genotype 225SF among free-ranging mule deer (Odocoileus hemionus) with chronic wasting disease. J Gen Virol 86:2127–2134. https://doi.org/10.1099/vir.0.81077-0
- Jiang Z, Yu C, Feng Z, Zhang L, Xia J, Ding Y, Lindsay N (2000) Reintroduction and recovery of Père David's deer in China. Wildl Soc Bull 28:681–687
- Johnson C, Johnson J, Clayton M, McKenzie D, Aiken J (2003) Prion protein gene heterogeneity in free-ranging white-tailed deer within the chronic wasting disease affected region of Wisconsin. J Wildl Dis 39:576–581. https://doi.org/10.7589/0090-3558-39.3.576
- Jones FW (1951) A contribution to the history and anatomy of Père David's Deer (Elaphurus davidianus). Proc Zool Soc Lond 121:319–370. https://doi.org/10.1111/j.1096-3642.1951.tb00800.x
- Kaluz S, Kaluzova M, Flint APF (1997) Sequencing analysis of prion genes from red deer and camel. Gene 199:283–286. https://doi.org/10.1016/S0378-1119(97)00382-X
- Kelly AC, Mateus-Pinilla NE, Diffendorfer J, Jewell E, Ruiz MO, Killefer J, Shelton P, Beissel T, Novakofski J (2008) Prion sequence polymorphisms and chronic wasting disease resistance in Illinois white-tailed deer (Odocoileus virginianus). Prion 2:28–36. https://doi.org/10.4161/pri.2.1.6321
- Klein J, Sato A, Nagl S, O'hUigín C (1998) Molecular trans-species polymorphism. Annu Rev Ecol Evol Syst 29:1–21. https://doi.org/10.1146/annurev.ecolsys.29.1.1
- Koenig D, Hagmann J, Li R, Bemm F, Slotte T, Neuffer B, Wright SI, Weigel D (2019) Long-term balancing selection drives evolution of immunity genes in Capsella. eLife 8:e43606. https://doi.org/10.7554/eLife.43606
- Kramm C, Gomez-Gutierrez R, Soto C, Telling G, Nichols T, Morales R (2020) In vitro detection of chronic wasting disease (CWD) prions in semen and reproductive tissues of white tailed deer bucks (Odocoileus virginianus). PLoS One 14:e0226560. https://doi.org/10.1371/journal.pone.0226560
- Kumar S, Stecher G, Li M, Knyaz C, Tamura K (2018) MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol Biol Evol 35:1547–1549. https://doi.org/10.1093/molbev/msy096
- Kurt TD, Sigurdson CJ (2016) Cross-species transmission of CWD prions. Prion 10:83–91. https://doi.org/10.1080/19336896.2015.1118603
- Kuznetsova A, Cullingham C, McKenzie D, Aiken JM (2018) Soil humic acids degrade CWD prions and reduce infectivity. PLoS pathogens 14:e1007414–e1007414. https://doi.org/10.1371/journal.ppat.1007414
- Kuznetsova A, McKenzie D, Banser P, Siddique T, Aiken JM (2014) Potential role of soil properties in the spread of CWD in western Canada. Prion 8:92–99. https://doi.org/10.4161/pri.28467
- Leigh J, Bryant D (2015) PopART: full-feature software for haplotype network construction. Methods Ecol Evol 6:1110–1116. https://doi.org/10.1111/2041-210x.12410
- Li C, Yang X, Ding Y, Zhang L, Fang H, Tang S, Jiang Z (2011) Do Père David's deer lose memories of their ancestral predators? PLoS One 6:e23623. https://doi.org/10.1371/journal.pone.0023623
- Librado P, Rozas J (2009) DnaSP v5: a software for comprehensive analysis of DNA polymorphism data. Bioinformatics 25:1451–1452. https://doi.org/10.1093/bioinformatics/btp187
- Manjerovic MB, Green ML, Mateus-Pinilla N, Novakofski J (2014) The importance of localized culling in stabilizing chronic wasting disease prevalence in white-tailed deer populations. Prev Vet Med 113:139–145. https://doi.org/10.1016/j.prevetmed.2013.09.011
- Mateus-Pinilla N, Weng H-Y, Ruiz MO, Shelton P, Novakofski J (2013) Evaluation of a wild white-tailed deer population management program for controlling chronic wasting disease in Illinois, 2003–2008. Prev Vet Med 110:541–548. https://doi.org/10.1016/j.prevetmed.2013.03.002
- Mathiason CK, Hays SA, Powers J, Hayes-Klug J, Langenberg J, Dahmes SJ, Osborn DA, Miller KV, Warren RJ, Mason GL, Hoover EA (2009) Infectious prions in pre-clinical deer and transmission of chronic wasting disease solely by environmental exposure. PLoS One 4:e5916–e5916. https://doi.org/10.1371/journal.pone.0005916
- Mathiason CK, Powers JG, Dahmes SJ, Osborn DA, Miller KV, Warren RJ, Mason GL, Hays SA, Hayes-Klug J, Seelig DM, Wild MA, Wolfe LL, Spraker TR, Miller MW, Sigurdson CJ, Telling GC, Hoover EA (2006) Infectious prions in the saliva and blood of deer with chronic wasting disease. Science 314:133–136. https://doi.org/10.1126/science.1132661
- Mead S, Stumpf MPH, Whitfield J, Beck JA, Poulter M, Campbell T, Uphill JB, Goldstein D, Alpers M, Fisher EMC, Collinge J (2003) Balancing selection at the prion protein gene consistent with prehistoric kurulike epidemics. Science 300:640–643. https://doi.org/10.1126/science.1083320
- Miller MW, Williams ES, Hobbs NT, Wolfe LL (2004) Environmental sources of prion transmission in mule deer. Emerg Infect Dis 10:1003–1006. https://doi.org/10.3201/eid1006.040010
- Mitchell GB, Sigurdson CJ, O’Rourke KI, Algire J, Harrington NP, Walther I, Spraker TR, Balachandran A (2012) Experimental oral transmission of chronic wasting disease to reindeer (Rangifer tarandus tarandus). PLoS One 7:e39055. https://doi.org/10.1371/journal.pone.0039055
- Monello RJ, Galloway NL, Powers JG, Madsen-Bouterse SA, Edwards WH, Wood ME, O’Rourke KI, Wild MA (2017) Pathogen-mediated selection in free-ranging elk populations infected by chronic wasting disease. PNAS 114:12208. https://doi.org/10.1073/pnas.1707807114
- Moore SJ, Kunkle R, Greenlee MHW, Nicholson E, Richt J, Hamir A, Waters WR, Greenlee J (2016) Horizontal transmission of chronic wasting disease in reindeer. Emerg Infect Dis 22:2142–2145. https://doi.org/10.3201/eid2212.160635
- Nichols TA, Pulford B, Wyckoff AC, Meyerett C, Michel B, Gertig K, Hoover EA, Jewell JE, Telling GC, Zabel MD (2009) Detection of protease-resistant cervid prion protein in water from a CWD-endemic area. Prion 3:171–183. https://doi.org/10.4161/pri.3.3.9819
- O'Rourke KI, Spraker TR, Hamburg LK, Besser TE, Brayton KA, Knowles DP (2004) Polymorphisms in the prion precursor functional gene but not the pseudogene are associated with susceptibility to chronic wasting disease in white-tailed deer. J Gen Virol 85:1339–1346. https://doi.org/10.1099/vir.0.79785-0
- Ohmura T, Ueda T, Hashimoto Y, Imoto T (2001) Tolerance of point substitution of methionine for isoleucine in hen egg white lysozyme. Protein Eng Des Sel 14:421–425. https://doi.org/10.1093/protein/14.6.421
- Pitra C, Fickel J, Meijaard E, Groves C (2004) Evolution and phylogeny of old world deer. Mol Phylogenet Evol 33:880–895. https://doi.org/10.1016/j.ympev.2004.07.013
- Pritzkow S, Morales R, Lyon A, Concha-Marambio L, Urayama A, Soto C (2018) Efficient prion disease transmission through common environmental materials. J Biol Chem 293:3363–3373. https://doi.org/10.1074/jbc.M117.810747
- Prusiner S (1991) Molecular biology of prion diseases. Science 252:1515–1522. https://doi.org/10.1126/science.1675487
- Prusiner SB (1982) Novel proteinaceous infectious particles cause scrapie. Science 216:136–144. https://doi.org/10.1126/science.6801762
- Raymond GJ, Bossers A, Raymond LD, O'Rourke KI, McHolland LE, Bryant PK 3rd, Miller MW, Williams ES, Smits M, Caughey B (2000) Evidence of a molecular barrier limiting susceptibility of humans, cattle and sheep to chronic wasting disease. EMBO J 19:4425–4430. https://doi.org/10.1093/emboj/19.17.4425
- Rhyan J, Miller M, Spraker T, McCollum M, Nol P, Wolfe L, Davis T, Creekmore L, O'Rourke K (2011) Failure of fallow deer (Dama dama) to develop chronic wasting disease when exposed to a contaminated environment and infected mule deer (Odocoileus hemionus). J Wildl Dis 47:739–744. https://doi.org/10.7589/0090-3558-47.3.739
- Richards B (2020) Expanding distribution of chronic wasting disease. U.S. Geological Survey. https://www.usgs.gov/centers/nwhc/science/expanding-distribution-chronic-wasting-disease?qt-science_center_objects=0#qt-science_center_objects
- Rivera NA, Brandt AL, Novakofski JE, Mateus-Pinilla NE (2019) Chronic wasting disease in cervids: prevalence, impact and management strategies. Vet Med (Auckl) 10:123–139. https://doi.org/10.2147/VMRR.S197404
- Robinson AL, Williamson H, Güere ME, Tharaldsen H, Baker K, Smith SL, Pérez-Espona S, Krojerová-Prokešová J, Pemberton JM, Goldmann W, Houston F (2019) Variation in the prion protein gene (PRNP) sequence of wild deer in Great Britain and mainland Europe. Vet Res 50:59–59. https://doi.org/10.1186/s13567-019-0675-6
- Saunders SE, Bartelt-Hunt SL, Bartz JC (2012) Occurrence, transmission, and zoonotic potential of chronic wasting disease. Emerg Infect Dis 18:369–376. https://doi.org/10.3201/eid1803.110685
- Schafer EH (1968) Hunting parks and animal enclosures in ancient China. JESHO 11:318. https://doi.org/10.1163/156852068X00126
- Slate J (2005) Molecular evolution of the sheep prion protein gene. Proc Biol Sci 272:2371–2377. https://doi.org/10.1098/rspb.2005.3259
- Stephens M, Smith NJ, Donnelly P (2001) A new statistical method for haplotype reconstruction from population data. Am J Hum Genet 68:978–989. https://doi.org/10.1086/319501
- Viggers K, Lindenmayer D, Spratt D (1993) The importance of disease in reintroduction programmes. Wildl Res 20:687–698. https://doi.org/10.1071/WR9930687
- Wildt D, Miller P, Koepfli K-P, Pukazhenthi B, Palfrey K, Livingston G, Beetem D, Shurter S, Gregory J, Takács M, Snodgrass K (2019) Breeding centers, private ranches, and genomics for creating sustainable wildlife populations. Bioscience 69:928–943. https://doi.org/10.1093/biosci/biz091
- Williams ES, Young S (1980) Chronic wasting disease of captive mule deer: A spongiform encephalopathy. J Wildl Dis 16:89–98. https://doi.org/10.7589/0090-3558-16.1.89
- Williams ES, Young S (1982) Spongiform encephalopathy of Rocky Mountain elk. J Wildl Dis 18:465–471. https://doi.org/10.7589/0090-3558-18.4.465
- Wulf M-A, Senatore A, Aguzzi A (2017) The biological function of the cellular prion protein: an update. BMC Biol 15:34–34. https://doi.org/10.1186/s12915-017-0375-5
- Yuan B, Xie S, Liu B, Xue D, Sun D (2019) Differential movement pattern of Père David's deer associated with the temporal rhythm using GPS collar fix. Glob Ecol Conserv 18:e00641. https://doi.org/10.1016/j.gecco.2019.e00641
- Zeng Y, Jiang Z, Li C (2007) Genetic variability in relocated Père David’s deer (Elaphurus davidianus) populations—implications to reintroduction program. Conserv Genet 8:1051–1059. https://doi.org/10.1007/s10592-006-9256-0
- Zhang C, Chen L, Zhou Y, Wang K, Chemnick LG, Ryder OA, Wang W, Zhang G, Qiu Q (2017) Draft genome of the milu (Elaphurus davidianus). GigaScience 7. https://doi.org/10.1093/gigascience/gix130
- Zhang X, Deng C, Ding J, Ren Y, Zhao X, Qin S, Zhu S, Wang Z, Chai X, Huang H, Ding Y, Lu G, Zhu L (2016) Comparative genomics and metagenomics analyses of endangered Père David’s deer (Elaphurus davidianus) provide insights into population recovery. bioRxiv:073528. https://doi.org/10.1101/073528
- Zhu L, Deng C, Zhao X, Ding J, Huang H, Zhu S, Wang Z, Qin S, Ding Y, Lu G, Yang Z (2018) Endangered Père David’s deer genome provides insights into population recovering. Evol Appl 11:2040–2053. https://doi.org/10.1111/eva.12705
Due to technical limitations, table 1 & 2 is only available as a download in the Supplemental Files section.
- SupplementaryFigure1.png
Supplementary Figure 1: Contiguous map of the United States showing in red the counties from which chronic wasting disease has been reported by the CDC in free-ranging cervids as of January 2020 (CDC 2020). The black icons indicate the locations of the institutions that provided Pere David’s deer samples for this publication: San Diego Zoo Institute for Conservation Research in San Diego County, California; Wildlife Conservation Society/Bronx Zoo in Bronx County, New York; and the Wilds in Guernsey County, Ohio. The blue icons indicate the locations of additional facilities that have Pere David’s deer in their collections. These include the Irving Game Farm in Hancock County, Indiana; Lake Superior Zoological Gardens in St. Louis County, Minnesota; the Lee G. Simmons Conservation Park and Wildlife Safari, in Saunders County, Nebraska; Memphis Zoological Garden & Aquarium in Shelby County, Tennessee; Oklahoma City Zoological Park in Oklahoma County, Oklahoma; Omaha's Henry Doorly Zoo in Douglas County, Nebraska; Virginia Safari Park in Rockbridge County, Virginia and White Oak Conservation Center in Nassau County, Florida. While institutions may not be in counties that have reported cases of chronic wasting disease (CWD), feed and bedding from regions with active cases of CWD pose a strong risk to the deer in captive breeding facilities (Henderson et al. 2015; Mathiason et al. 2009). Map made using ArcGIS v10.7.1.
- PerrinStowePereDavidsCWDTablesConGenSubmission.xlsx
{kind=link}