Low prevalence reported in this study by microscopic screening (Table 1) is not surprising considering the low sensitivity imminent of parasitological diagnostic method (28, 29, 30). This is especially so for field animals characterized by low parasitaemia. The superiority of PCR over MHCT have been widely demonstrated in the epidemiological study of animal trypanosomosis (3, 20, 32, 33). These differences are due to sensitivity thresholds of the techniques. As against the prevalence by microscopy, nested Polymerase Chain Reaction (PCR) method gave an overall prevalence of 3.02%. Each species tested produced amplicons of between 200–700 bp in length (Figs. 1 and 2). The ITS-1 PCR product size of T. evansi was similar to that of T. brucei and sequencing analysis was key to differentiating between the two PCR products. Sample JT11 was further confirmed to be T. evansi (supplementary data) suggesting the possible role of cattle as reservoirs of T. evansi. Generally, bands obtained from the amplification result were in agreement with previous studies (12, 31, 34, 35, 36, 38, 39, 40, 41). Samples JH4 and JM8 resulted in band sizes of approximately 700bp and which were confirmed by Sequence analysis to be Trypanosoma congolense Savannah sub-specie. Molecular characterization of Trypanosoma species using ITS-1 generic primers and/or its slight modification gave an estimated range of ITS-1 band sizes with a maximum amplicon length of 640bp (12) but went further in noting that all species amplification using generic primers could lead to a bands size of between 150–750bp in length as evidenced in this study and those previously reported (17, 31, 39, 42). An amplicon length of 210bp for T.vivax was not reported previously which may be an indication that T. vivax’s 18 rRNA is fast evolving at 7 to 10 times the rate of non-salivarian trypanosomes and also significantly evolving faster than all other trypanosomes. Although, no living specimen of this trypanosome was isolated, our conclusion could only be based on DNA sequence analysis while its taxonomic relationships were deduced from phylogenetic analysis of the amplified ssuRNA (70). Further biological characterization will depend on isolation of a living specimen into culture. The ability to identify this trypanosome by the distinct size of the ITS-1 region has provided preliminary information on its distribution and prevalence that should help track it down in the field. To make an evolutionary inference, all isolates aligned with the salivarian group except JO6 (Trypanosoma theileri) which fell in the stercoraria group. ssrRNA from JT11 (T. evansi), JAD7 (T. brucei), JAG2 (T. brucei) fell in the same branch but different clades which depicts a logical evolutionary event emanating from within these species (Fig. 3). Trypanosoma evansi is widely known to have evolved from Trypanosoma brucei and all of which were rooted on Trypanosoma vivax (16).
A drastic fall in PCV is traditionally considered a warning sign of the trypanosomiasis (43, 44). Classically, infection with trypanosome species that are pathogenic in local breeds of cattle result in retarded growth and anemia while nutritional status is a determining factor of infection (45, 46). From our findings, there is a PCV declined in trypanosome-positive cattle (Table 2) possibly due to the effect of parasites on blood cells. Similarly, the average hematocrit values varied between cattle breeds. However, the very low PCV presented by the red bororo breed may not be a true reflection of the PCV trend as only one animal was screened positive. The type of trypanosome specie infection impacted differently on the PCV of animals with average falling below the standard obtainable for cattle (24%-46%) except for the animals infected with Trypanosoma brucei with an average Packed Cell Volume of 30.3 ± 0.92. The animal infected with Trypanosoma theileri, a non-pathogenic trypanosome of cattle had the lowest PCV value (19.2 ± 1.12) (Table 3). From our findings, it may be illogical to conclude that this comparative decrease in the PCV is due to T. theileri infection as only one animal was infected. However, it is possible that the parasite may have transited from a non-pathogenic to pathogenic form, hence the need to have a controlled experiment aimed at monitoring the PCV in the face of Trypanosoma theileri infection and other trypanosome species.
This study showed 3.02% overall prevalence which compares well with 4.3% national prevalence as reported by European Economic Commission project of 1989 and 1996 (47), 3.9% in Ogbomosho (48), 4.69% in Oyo (49) and 9.4% in Kaduna (50) as against high prevalence of 53.4% in Kaura, Kaduna (51) and 46.8% in Jos (52). These contradictory findings might reflect seasonal or local differences in tsetse populations, sample size and site, improved sensitization among nomads on grazing course, better implementation on the use of trypanocidal drugs and urbanization which may have perturbed the ecology of the transmitting vector, leading to ecological migration to a more favourable ecosystem, hence low prevalence recorded in our study.
Although there is no any significant difference in the infection rate between male and female animals (Table 4), our results showed that females were more infected. This observed differences may be attributed to livestock management adopted in the farming community where larger numbers of males are frequently sold off the herd at any early age while the rest are kept for breeding or animal traction. Also female animals persist longer in herds for the purpose of breeding, thus allowing the chronic infection to be maintained for very long period. As a result, the remaining males are more closely monitored while the females are readily exposed to hazard in the population vis a vis multiple copulation with limited male animals in the herd. Also the larger population of females (59%) obtained in this study by simple random sampling may account for this difference. Previously, 199 male cattle and 121 female were examined with no statistical difference in the infection prevalence (49). However, occurrence of any disease is dependent on many factors of which sex is just one of them. Factors other than sex relating to the host or its environment could therefore have played a role in influencing the susceptibility of animals to infection which has been documented in several studies (8, 10, 53).
From our study, there is a decreases in disease prevalence as animals get older probably due to age-acquired immunity which could represent a key positive factor and bearing in mind that trypanocidal treatments are more frequently used on adults by local farmers. In addition, young animals are more vulnerable to tsetse bites due to their skin fragility. Moreover, they are not agile enough to ward-off insects away along the grazing route as the adults. The tsetse flies also frequently target weak animals as a source of food in order to avoid being crushed by moving animals (54). In this study, despite the very low number of young animals randomly sampled, prevalence of 10.0% (7/70) for younger animals and 1.5% (5/328) were recorded for adult which is statistically significant (P < 0.05) (Table 4). This indicated that the incidence rate was not similar in young and adult animals (8). Although not significant, the infection rate differ among cattle breed. The prevalence of trypanosome infection was lowest in Sokoto gudali (2.3%), a breed not known for trypanotolerance (56, 57), and may have resulted from adaptability of this breed to its environment. The higher prevalence observed among the White Fulani breed may be attributed to their trypano-susceptibility and perhaps due to their higher representation in the sampling (52.8%). Of the four cattle breeds studied, the White Fulani are usually raised under the nomadic system of management. This may be another possible explanation for the higher prevalence recorded by this cattle group (10).
Although our test of statistic showed no significant difference in the infection rate among cattle in regards to period of residency, the infection was more prevalent among animals that were recently domiciled (3.18%). This observation may well be attributed to recent influx of herders down south due to insecurity and ban on open grazing in some parts of Nigeria and which has forced nomadism away from the north. Similarly, our findings revealed that infection was found to be more prevalent among transhumance animals (3.6%) probably due to exposure to tsetse bites while pervading territories of different endemic locals in a bid to having greener pastures. Sentinel animals appear to be more protected from tsetse bites due to livestock management style and guided path to grazing by cattle keepers. Despite the difference in infection rate, the statistical test showed that the observed difference was due to mere probabilistic chance (Table 4).
Having excluded animals with recent administration of trypanocidal drugs, a high prevalence of the disease was noted among animals having disease history (4.05%). An explanation to this could be that animals having the disease history may have not been well treated to clear the parasite in their blood in the first place or it may be that the treatment administered during the last infection may not be effective/efficacious or the parasite itself may have developed resistance to the administered drugs. During our survey, 92% of sampled animal had history of trypanocidal drugs treatment (Table 4) which may explain the low infection prevalence generally recorded with possible indications of drug resistance as seen in animals known to have had history of the disease and treatment but still reported in the study as infected animal.
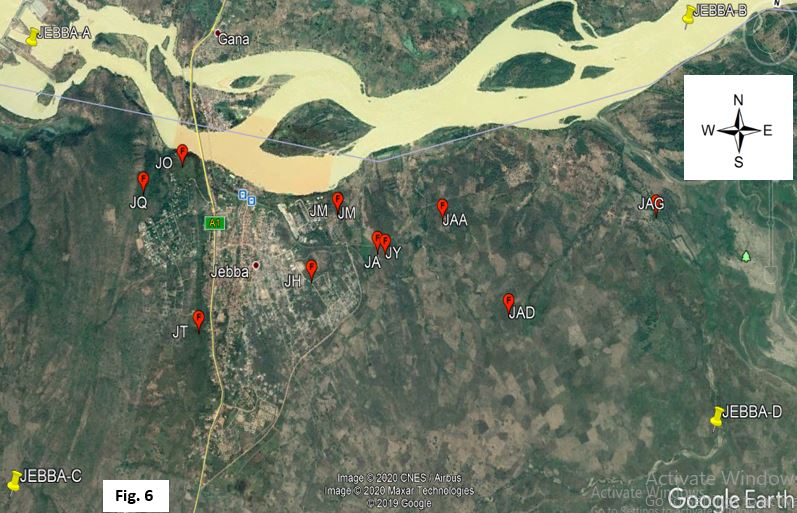
The prevalence of trypanosome infection was significantly higher in locations closer to river Niger as compared to those further away (Table 5 and Fig. 6). This may be attributed to differences in herd management practices, grazing route which predisposes the herd to tsetse bites, herd composition and frequent exposure to trypanocidal drugs which may differ in each herd. The river could be a positive factor for the vector transmitting the disease as well as a source of water for grazing animals which could expose them to risks of bite by riverine species of the flies (58).
Although not significant, the disease rate was high among larger herds (33.3%) as compared to smaller ones (23.8%) (Table 5). It may be that smaller herds are more closely monitored and easily managed and treated before the transmission sets in as compared to larger herds where animals are seen as a single entity. Furthermore, animals that had travel history to trypanosomiasis endemic zone of the country (Benue, Jos, Kaduna, Delta, Oyo and Ogun as published in literatures) were more infected (50.0%) probably due to contact of travelling animals with infected sentinels in endemic zones and exposure to tsetse bite during trans-boundary movement.
Majority of the trypanosomes in cattle were T. congolense and T. brucei which accounted for 50.0% (6/12) and 16.67% (2/12) (Fig. 5) respectively with nearly half of the overall infection due to Trypanosoma congolense Savannah sub-specie (Fig. 6), possibly as a result of large host range or probably due to the fact that riverine species of tsetse are generally considered susceptible to T. congolense infections (69). High prevalence of T. congolense infection is an indication of the dominance of G. mosrsitans species of the fly (7, 8, 18, 20, 52, 59, 60, 52) and could be that its transmission is highly favoured by the obligate cyclical vector or the T. vivax and T. brucei respond better to the trypanocidal drugs, diminazene aceturate and homidium chloride administered by farmers. A high prevalence of the Savannah subgroup in cattle may also indicate that the parasites were introduced recently into the tested herds coupled with its reported virulence as compared to other sub-specie (55). The low prevalence of T. brucei infection may relate to reported resistance of indigenous West African cattle to the parasite (61). However, the detection of T. brucei and T. evansi in Nigerian cattle might portend serious danger not only to cattle and other livestock but also to livestock owners and the host communities at large as T. evansi infection has been reported in cattle and humans in India (62, 63). Low prevalence of T. simiae infection is an indication of low transmission of the parasite as animals infected with this species will probably not survive the acute and severe nature of this parasite (64, 65). Double infections in animals are a normal occurrence in the field (66). This study identified T. congolense Kilifi and T.vivax mixed infection in only one of the herd clusters. In Nigeria, previous surveys identified mainly T. congolense and T. vivax as animal pathogenic trypanosomes (67) and co-circulation has been reported in studies conducted in northern Nigeria (20, 41, 68). Co-infections with multiple Trypanosoma species have also been documented previously due to bites from tsetse flies carrying more than one Trypanosoma infections or successive bites from flies with different Trypanosoma species (8, 16, 37).




{kind=link}