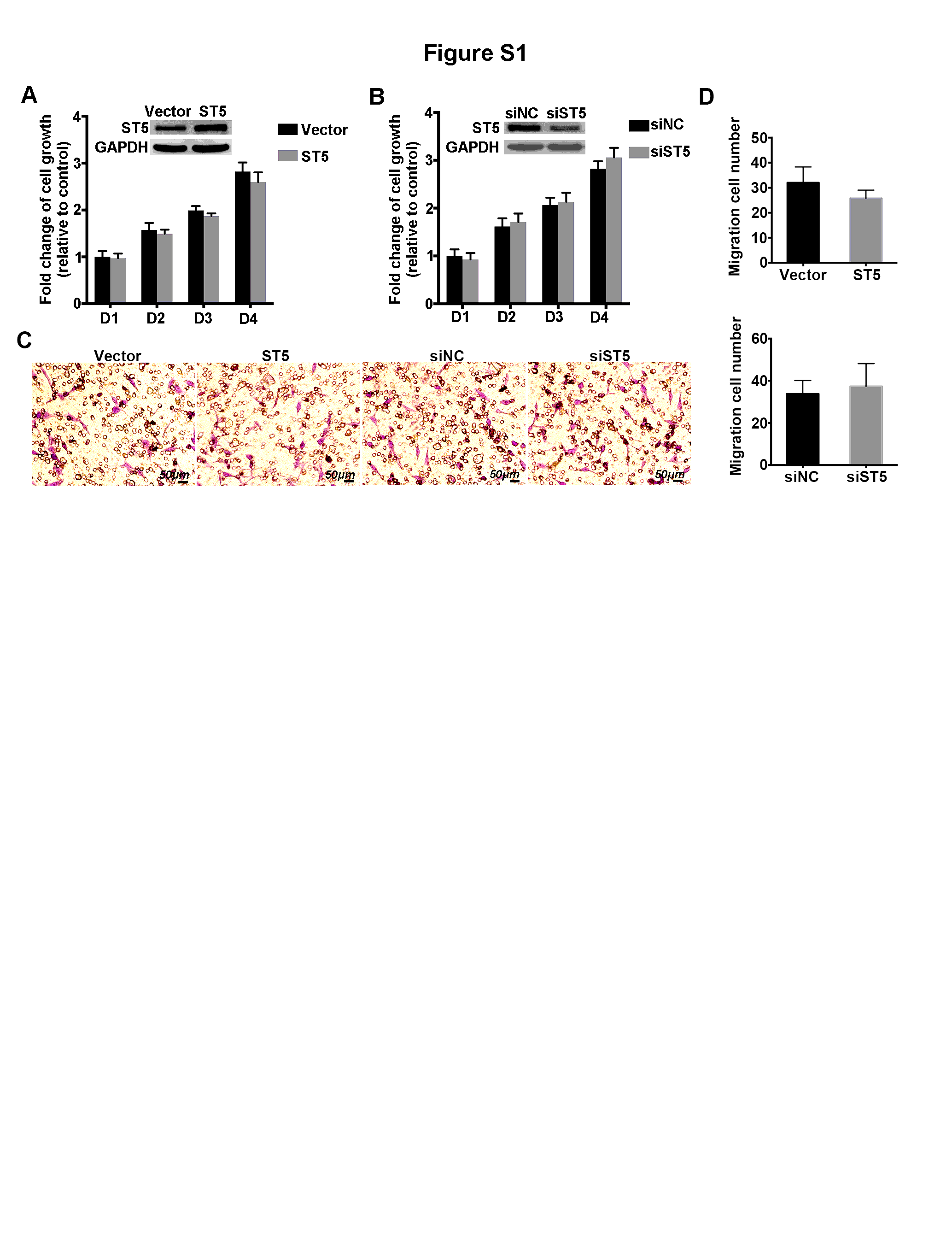
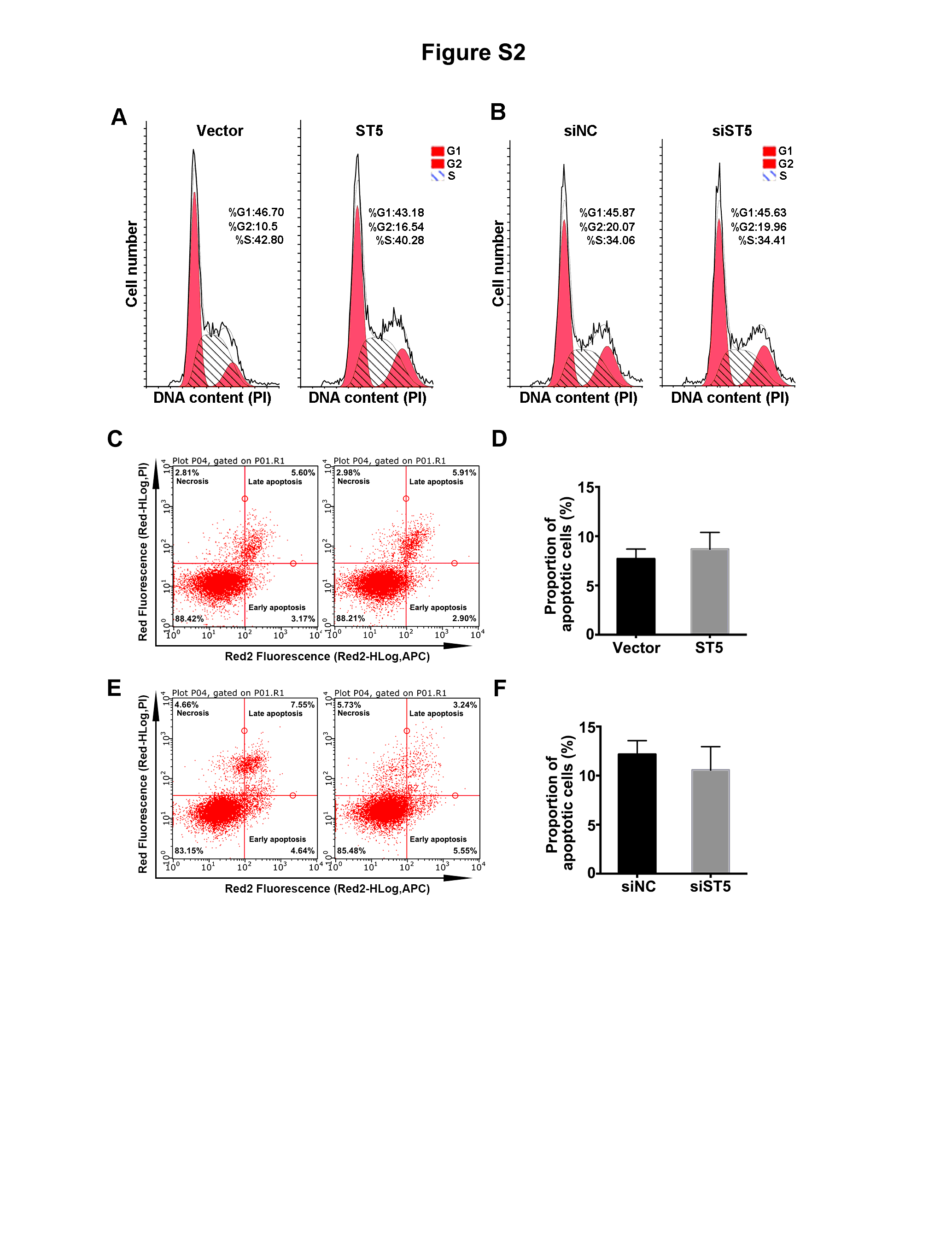
Currently, a combination of surgery and chemotherapy or radiation therapy is the most common treatment for breast cancer; however, poor prognosis and relapse still occur frequently in patients with distant metastases (13). Although advanced targeted cancer therapy and neoadjuvant therapy are constantly emerging, they are associated with several limitations for blocking cellular migration (14–17). ST5 shows differentially expression in tumorigenic and nontumorigenic somatic cell hybrids of Hela cells, implying ST5 might affect tumorigenesis (18). In the present study, ST5 upregulation abolished tumor migration of breast cancer cells and hypermethylated ST5 might implicated in this progression. Probably, ST5 acts as a potential therapeutic target for metastatic breast cancer.
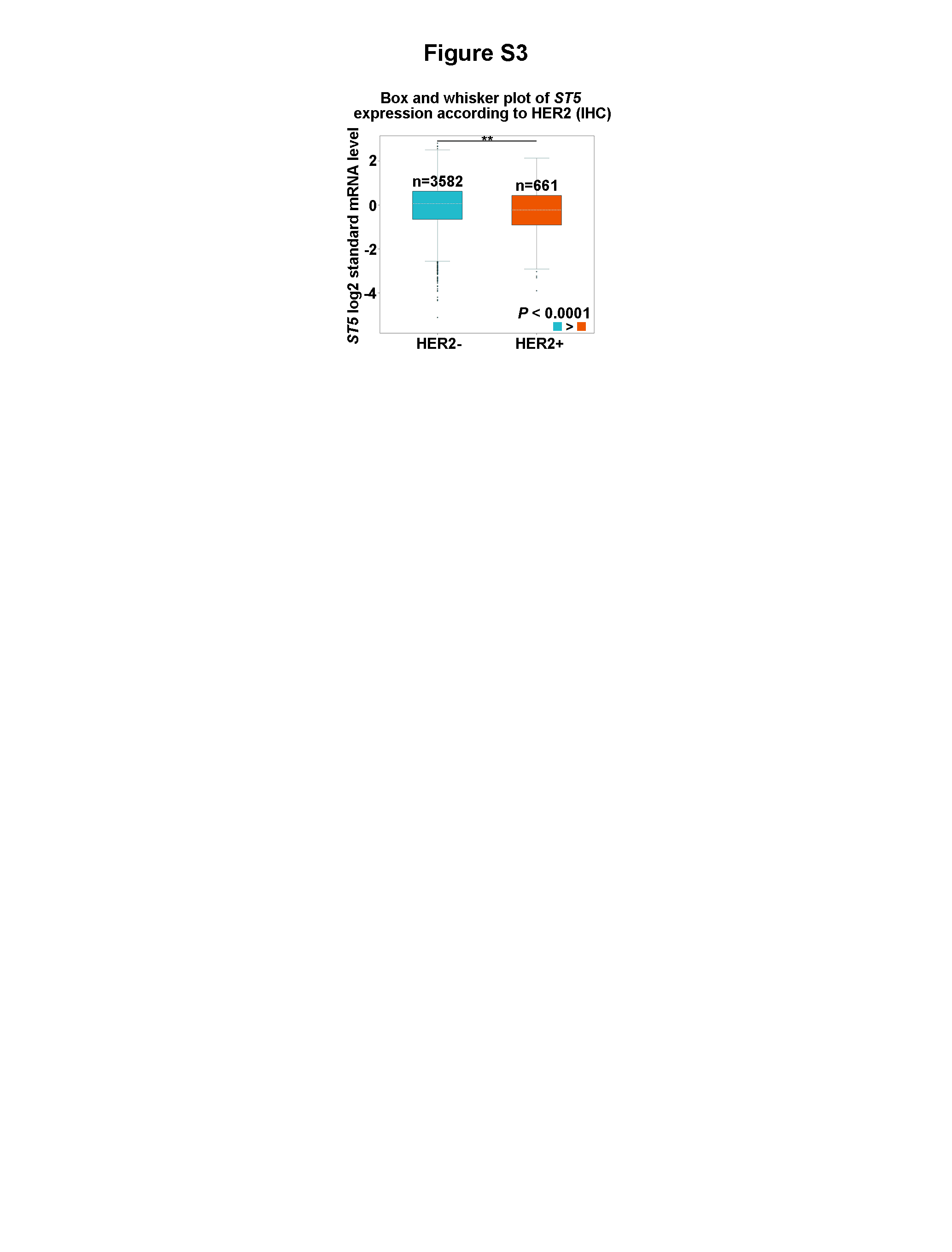
ST5 is differential expression in several gynecological oncology (8, 11). ST5 (P70kD) decreases tumorigenicity in HeLa cells, implying it may serve as a cancer suppressor gene (10, 19). Here, the decline of full length ST5 was observed in human breast cancer, particularly in invasive breast cancer cells. The finding firstly demonstrates ST5 (P126kD) shows differential expression among normal mammary epithelial cells, non-invasive and invasive breast cancer cells. However, the data does not detect the alterations of the other two variants among these cell types. Interestingly, the high invasive capability cell line SKBR3, which is ER- and progesterone receptor (PR)-negative but HER2-positive, also presented lower ST5 levels than TNBC cells (MDA-MB-231). On the basis of analysis results from bc-GenExMiner v4.4, ST5 status also appeared a significant decline in HER2-positive breast cancer patients (n = 661) compared to that in HER2-negative patients (n = 3582) (Fig. S3). The data imply a potential association between HER2 and ST5 in breast carcinogenesis which still needs to study further. Additionally, ST5 (P70kD) is correlated with decreased tumorigenic phenotype in mammalian cells, with a restoration in their transformed phenotype and contact-dependent growth (9, 20). Based on our present data, full length of ST5 (P126kD) also possessed the inhibition effect on biological behavior of invasive breast tumor cells, though previous study has indicated that ST5 (P126kD) promoted metastatic behavior of normal breast epithelial cell (12). Of note, in our data, we did not exclude the impact of ER that was expressed in MCF-7 cells, but not in MDA-MB-231 cells. Whether overexpression of ST5-mediated antitumor effect is prevented by the presence of ER in MCF-7 cells remains unclear.
ST5 is important for cellular signal cascades and the regulation of the transformed phenotype (21). Structurally, ST5 contains a group of GDP/GTP exchange region for Rab protein family and the mitogen-activated protein (MAP) kinase-activating death region which is verified as a interactant of TNF-alpha receptor (22, 23). ST5 can activate the MAP kinase ERK1/2 in response to epidermal growth factor (EGF) in cos-7 African green monkey fibroblast cells (24). The proline-rich region of ST5 isoform can recognize Src homology 3 (SH3) binding domains and MAP kinase phosphorylation sites (25–27), and participated in RAS/MEK2/ERK2 signaling cascade (20). Additionally, DENN/MADD, a multifunctional domain protein, interacts with JNK, activating MAPK/JNK pathway in Alzheimer's disease (AD) pathogenesis (28, 29). Therefore, ST5-mediated intracellular signaling transduction has the involvement of ERK1/2/JNK pathway. In the present study, we observed that ERK1/2/JNK activation was obviously inhibited, followed by the decrease of c-Myc in MDA-MB-231 cells transfected with exogenous ST5, vice versa. Possibly, ERK1/2/JNK signal axis was the downstream of ST5 during the migration process of breast tumor cells. However, whether ST5-mediated anti-tumor effect depends on ERK1/2/JNK pathway needs more investigation.
Low expression of tumor suppressors was regulated by diverse of factors, such as microRNAs, transcription factors, and DNA methylation (30, 31). Similar to other cancer types, human breast cancer also exhibits a high number of epigenetic alterations in the genome (32). Methylation profiles can be used as clinical biomarkers to predict drug response and prognosis of breast cancer (33, 34). DNA methylation alterations associated with cancer-related genes have become more pronounced and acted as the effective biomarker for early diagnosis and disease progression monitoring in the invasive breast tumors (35–37). The previous research from our group has revealed hypermethylated ST5 in gastric poorly differentiated adenocarcinoma (38). Herein, ST5 was also highly methylated in invasive breast cancer cells. And methylation inhibitor not only promoted ST5 expression, but also rescued tumor characteristics of MDA-MB-231 cells. Probably, the promoter methylation of ST5 regulated ST5 decline-evoked cell viability and migration enhancement.
{kind=link}
{kind=link}
{kind=link}