In this paper, we investigated the consequence of STAU2 depletion in non-transformed and cancer cells. We show that STAU2 depletion accelerates cell cycle progression in non-transformed cells but not in cancer cells. Interestingly, under physiological conditions, the steady state level of STAU2 protein is controlled by caspases and by the activity of the CHK1 pathway. This loop likely contributes to the fine-tuning of cell proliferation in changing cell environment. The regulation of STAU2 at the protein level is in addition to that already observed at the transcriptional level by the transcription factor E2F1 [25]. Regulation of STAU2 by the E2F1 and CHK1 pathways that induce and facilitate S phase progression and DNA surveillance suggests a role for STAU2 in pathways that merge with mechanisms of DNA replication and/or DNA maintenance during S phase.
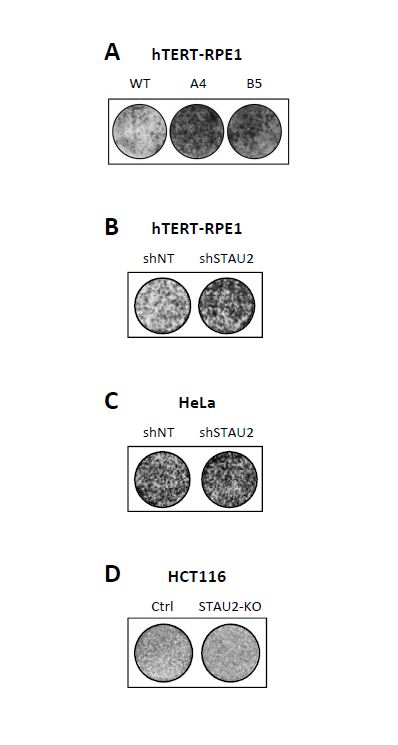
STAU2 regulates cell proliferation
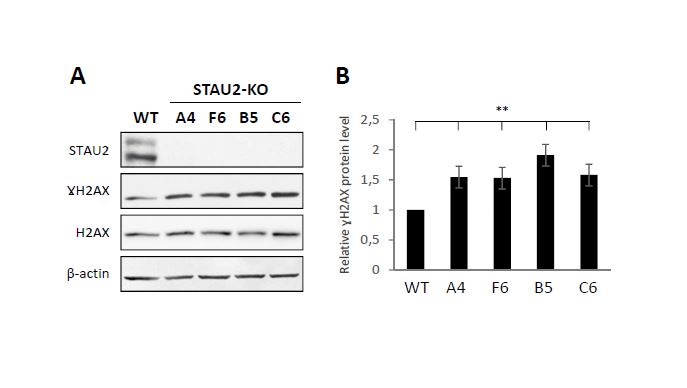
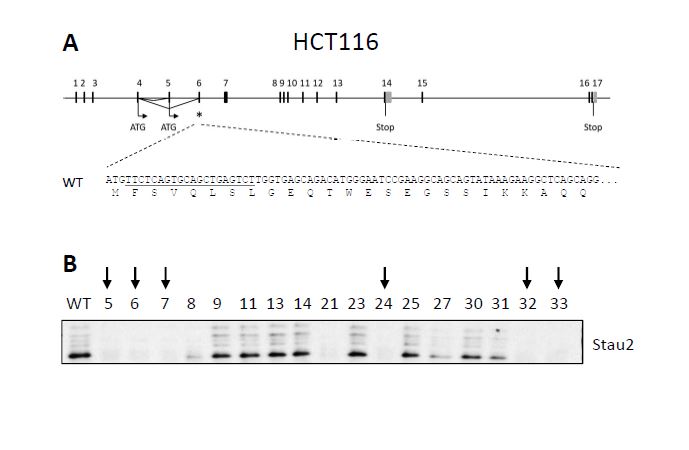
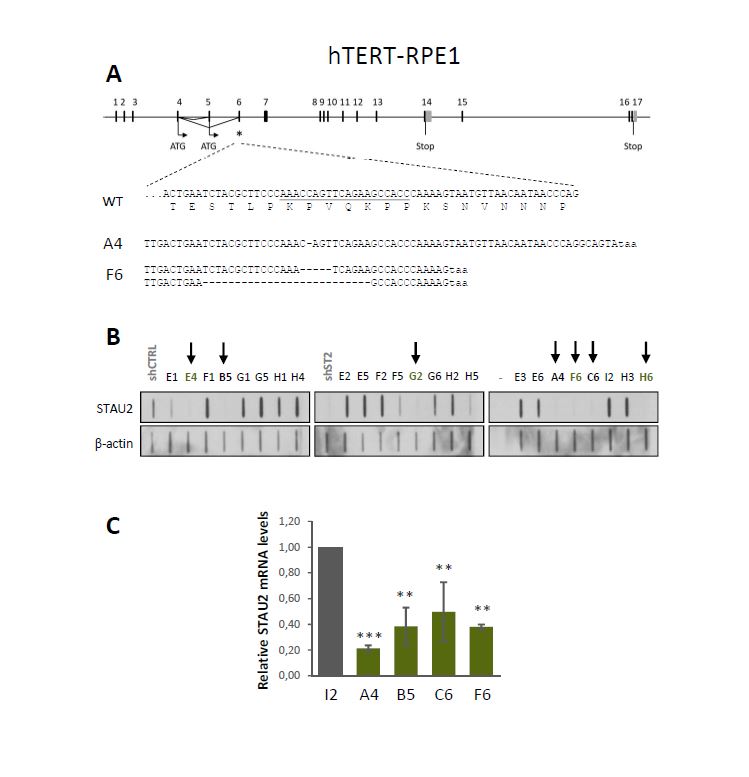
STAU2, as an RNA-binding protein involved in the posttranscriptional regulation of gene expression, was previously shown to bind mRNAs coding for proteins involved in cell cycle regulation and cell cycle proliferation [10], some of them having proto-oncogene or tumor suppressor functions. Notably, STAU2 was shown to bind 26 mRNAs involved in the TGF-β pathway, including the central protein TGF-β receptor I [10]. It was recently reported that dysregulation of the TGF-β pathway in RPE1 cells leads to cell cycle dysregulation and tumoral transformation [38]. The phenotypes observed upon STAU2 depletion are indeed compatible with oncogenic transformation. Oncogenic transformation was previously observed when RNA-binding proteins inappropriately misregulated proto-oncogenes and/or tumor suppressor genes [6, 39, 7, 40, 3]. The hyper-proliferative phenotype that results from the misregulation of these RNA-binding proteins is often followed by an accumulation of DNA damage due to the loss of cell cycle checkpoints [41, 42]. Accumulation of DNA damages are indeed observed in STAU2-KO hTERT-RPE1 cells (Additional Fig S4) and in RNAi-mediated STAU2 knockdown in HCT116 cells [25]. Oncogenic transformation may also explain why tumor cells are not affected by STAU2 depletion. Transformed cell lines are known to be permanently stimulated by oncogenic stimulus [43, 44], and therefore additional oncogenic stimulus may not affect their proliferation.
STAU2 protein is degraded by caspases and stabilized by the CHK1 pathway
Under physiological conditions, STAU2 protein level is decreased by caspase activity and stabilized by the CHK1 pathway. As CHK1 activation inhibits caspase activity [45], it is tempting to propose that CHK1 indirectly controls STAU2 steady-state levels via the activation/inhibition of caspase activity. Although well characterized for their roles in response to massive DNA damages and induction of apoptosis [46–50], caspases also play other important roles in unstressed cells related to cell proliferation, differentiation and cellular reprogramming [51–53]. In turn, CHK1 is involved in cell cycle control, especially during the S phase, where it is required for firing origin of DNA replication [54]. CHK1 regulates checkpoint controls and is essential for cell survival in dividing cells [55] and its inhibition induces an accumulation of DNA damage and apoptosis [56, 57].
Alternatively, CHK1 may control STAU2 stability via activation of downstream kinases as previously described for the CHK1-mediated control of the RNA-binding protein HuR via the regulation of CDK1 [58]. Interestingly, STAU2 is a target of CDK1 [30] and CDK1 was found in the list of proteins in proximity to STAU2 (Additional Table S1). In contrast, we have no evidence that STAU2 is a direct target of CHK1. CHK1 was not found in the list of proteins in proximity to STAU2 and, using an in-vitro kinase assay, we did not observe phosphorylation of STAU2 by a purified CHK1 kinase (data not shown).
STAU2 is in close proximity to proteins involved in RNA posttranscriptional regulation and proteins of the nucleolus
Degradation/stabilization of STAU2 and/or modulation of STAU2 functions via proximal partners should influence the pathway(s) in which STAU2 is involved. The BioID2 and TurboID experiments that detect proteins in close proximity to STAU2 link STAU2 to its well recognized functions in RNA posttranscriptional regulation. Indeed, STAU2 is well known for its roles in translation [15], RNA localization [12, 9, 15], splicing [14], mRNA decay [16], and stress granule formation [59]. It is likely that STAU2 regulation by caspases and/or the CHK1 pathway fine tunes the expression of mRNA regulons, resulting in well-ordered cell proliferation.
Interestingly, these genome-wide experiments also identify many proteins that can be found in the nucleolus. The nucleolus is well known for its role in ribosome biogenesis and for its involvement in non-ribosomal ribonucleoprotein complex formation [60]. Labeling of proteins found in the nucleolus compartment is consistent with earlier observation showing that STAU2 can migrate in the nucleolus [11]. The role of STAU2 in the nucleolus is unclear but it was proposed that STAU2 may assemble ribonucleoprotein complexes in the nucleolus [61]. The presence of ribosomes in STAU2-containing ribonucleoprotein complexes [9] further suggests a functional link between ribosome biogenesis and RNP formation in the nucleolus. STAU2 indeed co-immunoprecipitates with the ribosome biogenesis factor RSL1D1 [31]. The nucleolus might function as a checkpoint to verify the potential functional integrity of RNP and RNP-ribosome complexes or to tightly regulate their activity and/or release. Depletion of STAU2 may thus cause the formation of less-functional ribonucleoprotein complexes that could be released ahead of time or in inappropriate cellular compartments, impairing cell proliferation.
Other proteins in proximity to STAU2 are members of the DNA damage response. Most of these proteins can be found in the nucleoplasm and in the nucleolus and, therefore, could be non-specifically labeled via their co-localization with STAU2 in the nucleolus. However, STAU2 depletion causes an accumulation of DNA damages and therefore could be somehow involved in processes of DNA repair. Several processes could link STAU2 to DNA repair pathways. First, rDNA genes are the most transcribed genes and thus collisions between the transcription machinery and the replication fork are frequent [62, 63]. Consistently, several proteins involved in DNA replication are found in proximity to STAU2. The clashes between the machineries lead to replication fork stalling and single-stranded breaks [60]. This, in turn, activates the CHK1 kinase pathway [64–66]. In addition, R-Loops forms between nascent mRNAs and template DNA strands leading to genomic instability [67–69]. Remarkably, dysregulation of post-transcriptional processes increases R-loops formation [67, 70]. Depletion of STAU2 may thus facilitate the formation of R-loops whereas STAU2 stabilization upon CHK1 activation may help to attenuate the effects of R-loop formation. Alternatively, several studies have revealed the importance of lncRNAs in the DNA damage repair processes [71]. lncRNAs act as scaffolds for DNA repair protein recruitment. For example, lncRNA LINP1 participates in DNA-PK recruitment to DSB [72] and lncRNA NEAT1 is needed for the formation of DNA-PK/HEXIM1 paraspeckle complexes [35]. In other cases, lncRNAs play a role in chromatin modification, which is an indispensable step for the DNA repair process [73]. It is conceivable that STAU2 may be required to localize lncRNAs at the DNA damage sites to support the repair process.
{kind=link}
{kind=link}
{kind=link}
{kind=link}