Diet shifting and adaption affects phenotypic traits
Fish have the potential to change their behavioral and physiological characteristics, such as food selection, food intake, enzyme production, and development of the alimentary tract, to adjust to differences in the quality of available food [27, 29]. Generally, short-term adaptive changes are limited by the range of phenotypic plasticity, and long-term adaptive changes depend on the evolution of the genotype. Previous studies have indicated that differences in the histological structures of the fish gastrointestinal tract are related to feeding habits, food, age, and weight [30, 31]. In this study, the allodiploid hybrid fish that were adapted to carnivorous and herbivorous diets also showed a significant difference in histological structures of liver and intestinal tissues (Figure 1), indicating that food type participates in the developmental process of the gastrointestinal tract of the hybrid fish [14-15].
Genetic adaptation to dietary environments is a key process in the evolution of natural populations and is of great interest in fish breeding, as the ability to adapt to major dietary changes can be used effectively to promote fish welfare and more sustainable aquaculture. Recently studies have focused more on the effect of feeding changes (substituted diets) on the growth performance, metabolism, nutrition, and gene regulation of fishes [32-34]. However, dietary changes and adaptation are more or less affected by the function of hepatic metabolism, which may result in problems related to metabolism and other problems, such as reduced antioxidant capacity, disordered glucose and lipid metabolism, and weakened disease resistance [35-37]. In this study, due to the merge of subgenomes from the carnivorous TC and herbivorous BSB, the hybrid fish seems to have the potential to adapt to carnivorous, herbivorous, or omnivorous diets; in fact, no impact on growth level was observed between the two diet group of fish. Metabolites related to antioxidant protection (GSH, T-SOD, and POD) in the liver were not different between the two groups of fish. A higher concentration of MDA was detected in the carnivorous fish (Table 5), indicating that adaptation to a carnivorous diet affects hepatic metabolism in the hybrid fish; changes in metabolic enzyme activity need further elucidation.
Diet shifting and adaption affects metabolic pathways
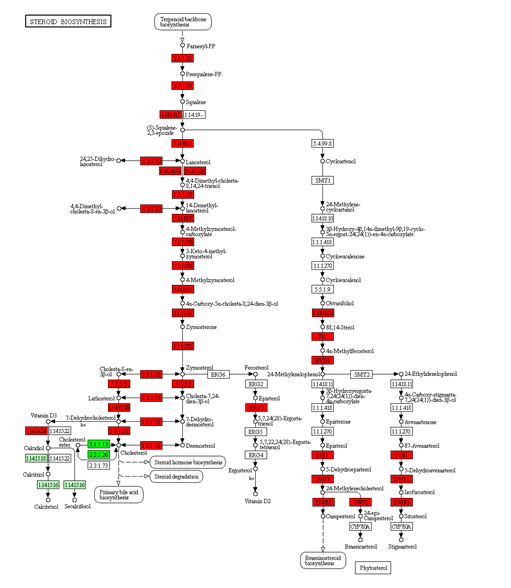
Diet shifting and long-term adaption can affect the digestion and metabolism of major nutrients (carbohydrates, proteins, and lipids) and often leads to significant metabolic changes (such as the modification of certain key metabolic enzymes and hepatic metabolic pathways, including energy metabolism, cholesterol biosynthesis, and proteolytic activity pathways) [35, 38-41]. In the current study, a number of DEHs were identified to be involved in the adaptation of hybrid fish to a herbivorous or carnivorous diet (Figure 2 and Table 2). The DEHs were highly enriched in carbon metabolism, steroid biosynthesis, and amino acid, fatty acid, and protein metabolism (Table 3, Figure S1 and S2), indicating that diet informs genetic adaption, which adjusts to available food. Metabolic adaption of these pathways apparently supports the adaptation of the hybrid fish to a carnivorous or herbivorous diet. In European sea bass (Dicentrarchus labrax), genes involved in the cholesterol biosynthetic pathways were upregulated when they were fed a plant diet compared to a fish-based diet [42]. In grass carp (Ctenopharyngodon idellus), a herbivorous fish, the genes involved in steroid biosynthesis, terpenoid backbone biosynthesis, and glycerophospholipid metabolic pathways were differentially expressed during the transition from a carnivorous to a herbivorous diet [43]. In BSB, feeding of a high fat-carbohydrate diet for 8 w also caused a significant decline in the number of amino acids entering the TCA cycle, which in turn resulted in elevated levels of seven amino acids [44].
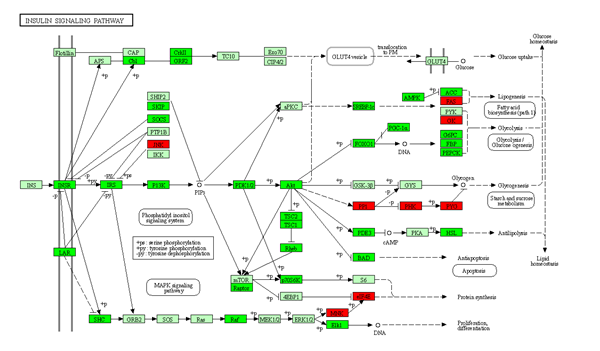
Insulin serves as the primary regulator of the blood glucose balance, regulating the metabolism and storage of nutrients, such as proteins, sugar, and fat, by acting on the liver, muscle, and adipose tissue [45]. In BSB, feeding of a high-carbohydrate diet for 8 w upregulated the genes associated with insulin signaling pathways, which could lead to the development of insulin resistance in hepatocytes, pathological liver changes, and, eventually, non-alcoholic, fatty liver disease [35]. The current study also identified that the insulin receptor, insulin receptor-related receptor, and downstream pathways, including PI(3)K, Akt, FOXO1, Cbl proto-oncogene, and mTOR pathways, were down-regulated in the herbivorous fish (Fig. S2), suggesting that the adaptation of hybrid fish to a carnivorous diet may also cause insulin unbalance and insulin resistance in hepatocytes [35, 45].
Homoeolog expression silence and bias contribute to dietary adaptation
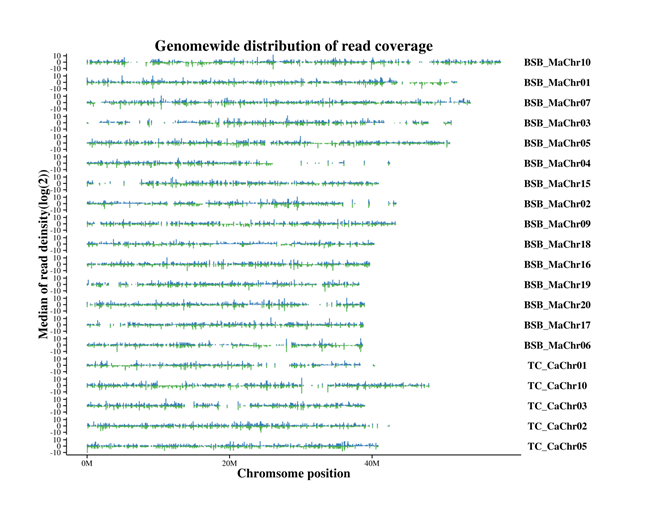
The merged subgenomes in a allopolyploid must adjust and coexist with one another in a single nucleus; this may cause interactions between diverged regulatory networks due to differential dosage sensitivities and epigenetic alterations. Variable expression of the homoeolog gene, resulting from such subgenome interaction, has been reported in some allopolyploid plants [9-13]. To some extent, investigating the variable expression of homeologous genes is hindered by at least two major obstacles: (1) lack of genomic resources for extant parental progenitors, and (2) the inability to distinguish the origin of the subgenome. Our previous study completed the whole genome sequencing of the two parents (BSB and TC) of the hybrid fish, providing basic data for the present study [28]. The parent subgenome was observed to be selectively expressed in tissues, and TC exerted a higher subgenome expression dominance than BSB (Figure 3). One possible explanation for this is that the newly formed hybrid must resolve genetic incompatibilities very rapidly within the first generation to adapt to the environment and facilitate reproduction [7, 9]. The subgenome expression dominance may be associated with inherent dominance between subgenomes. Silenced homoeolog genes were unbalanced in tissues, and the number of expression silenced genes was significantly more than the number of expression bias gene during dietary adaptation (Table 4). This may be associated with various regulatory elements, such as cis and trans regulatory elements, less transposable elements, and DNA methylation [46]. However, the mechanisms to preferentially silence one parental subgenome over other subgenomes and the functions of the subgenomes require further study.
How one subgenome is more expressed and whether the direction of subgenome dominance is externally induced in a polyploid is largely unknown. However, a possible way to resolve genetic incompatibilities is that some certain subset of pathways are controlled by one subgenome, and the rest pathways are controled by another subgenome [11, 47]. Obviously, this would result in the partitioning of phenotypic traits to different subgenomes, and it is of great significance in genetic breeding of plants and animals. Subgenome dominance contributing to heterosis (such as phenotypic bias) has been observed in cotton, wheat, and blueberry plants [9-12]. In the present study, the parent subgenome controlled functional pathways during dietary adaptation. The BSB subgenome mainly controlled the metabolism and diseases pathways, while the TC subgenome mainly controlled genetic information processing and cellular processes (Figure 4). Pathways, such as those involved in the circadian rhythm and digestion and metabolism pathways, were also activated in the grass carp during a transition from a carnivorous to herbivorous diet [43, 48]. Pathways involved in retinal photosensitivity, circadian rhythm, and appetite control show altered expression in mandarin fish (Sinipercidae) fed only live prey [49]. Subgenome expression dominance may play key roles in the adaptation of hybrid fish to herbivorous or carnivorous diets. The dynamic expression level of the homoeolog genes may provide a better understanding of the functional pathways that controlled by parent subgenome. However, a limiting factor is that the discontinuity of the SNP site makes it insufficient to design ideal primers. In addition, it will be interesting to investigate whether subgenome expression dominance is heritable and contributes to diet selection in hybrid F2-F5 generations.
{kind=link}
{kind=link}
{kind=link}