Effects of broccoli residues on VW of different cotton cultivars
Broccoli residues had a significant impact on the disease incidence and disease index of cotton VW (P༜0.05). Compared with the blank control (no broccoli residues), the disease incidence of cultivar EJ-1 decreased by 38.76% and 53.50% and the disease index decreased by 46.47% and 57.04% in Field A and Field B, respectively. The disease incidence of cultivar J863 decreased by 100% and 63.42% and the disease index decreased by 100% and 72.30% in Field A and Field B, respectively. The average control effects for EJ-1 and J863 were 51.76% and 86.15%, respectively (Fig. 1).
Effect of broccoli residues on DNA copies of V. dahliae in soil
When compared with those in the blank control soils, the DNA copies of V. dahliae in the soils associated with the different cotton cultivars were significantly reduced by the BR treatment (Fig. 2). For EJ-1, the DNA copies of V. dahliae decreased by 14.31% and 23.44% in Field A and Field B, respectively. For J863, the DNA copies of V. dahliae decreased by 34.19% and 26.34%, respectively. The average numbers of DNA copies of V. dahliae for EJ-1 and J863 were decreased by 18.88% and 30.27% following the application of BR, respectively.
Alpha Diversity Of The Bacterial Community
The alpha diversity of the bacterial community was expressed by the ACE and Chao1 indices in our study (Fig. 3). In Field A, the ACE index for EJ-1 ranged from 2503 (CK) to 2667 (BR), and the Chao1 index ranged from 2501 (CK) to 2624 (BR), which were greater by 6.55% and 4.92%, respectively. The ACE index for J863 ranged from 2603 (CK) to 2652 (BR), and the Chao1 index ranged from 2585 (CK) to 2617 (BR), which were greater by 1.89% and 1.24%, respectively. In Field B, the ACE index for EJ-1 ranged from 3690 (CK) to 3751 (BR), and the Chao1 index ranged from 3466 (CK) to 3541 (BR), which were greater by 1.64% and 2.15%, respectively. The ACE index for J863 ranged from 3949 (CK) to 3972 (BR), and the Chao1 index ranged from 3520 (CK) to 3655 (BR), which were greater by 0.59% and 3.85%, respectively. These results indicate that the ACE and Chao1 indices were increased by the application of BR at the different field sites.
Bacterial Community Structure Analyses
Principal component analysis based on the OTU composition was used to study the effect of broccoli residues on the soil bacterial community structure associated with the different cotton cultivars. Figure 4 shows plots of the sites in the plane of the first two principal coordinates based on the soil bacterial communities in Field A and Field B, respectively. The results show that the bacterial community structure associated with the different cultivars was located in the same quadrant after the application of broccoli residues, while that of the blank controls of the different cultivars was located in different quadrants, which indicates that the bacterial community structure changed and tended to be the same after the application of broccoli residues. In addition, the first principal component (PC1) and the second principal component (PC2) of the bacterial community structure at the OTU level in rhizosphere soil were found to explain 34.07% and 16.36% of all variables in Field A, and 24.83% and 21.29% of all variables in Field B, respectively. The cumulative contribution rates of variance of the two principal components reached 50.43% and 46.12%, respectively. In addition, ANOSIM indicated that the BR treatment contributed significantly to the separation of the CK treatment (R = 0.9815, P = 0.001, Field A) and (R = 0.6481, P = 0.002, Field B).
Comparison Of Bacterial Community Composition
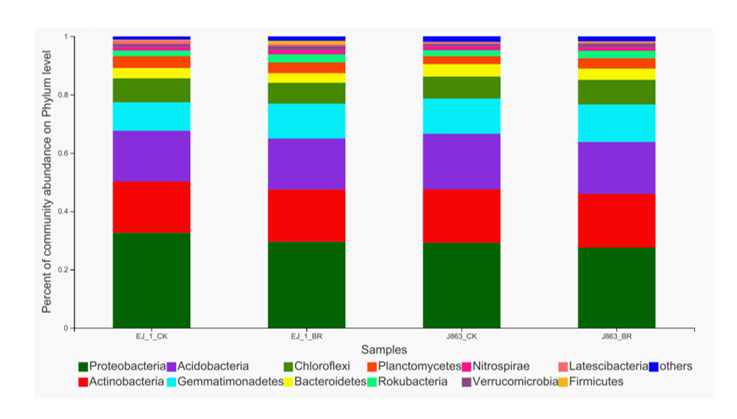
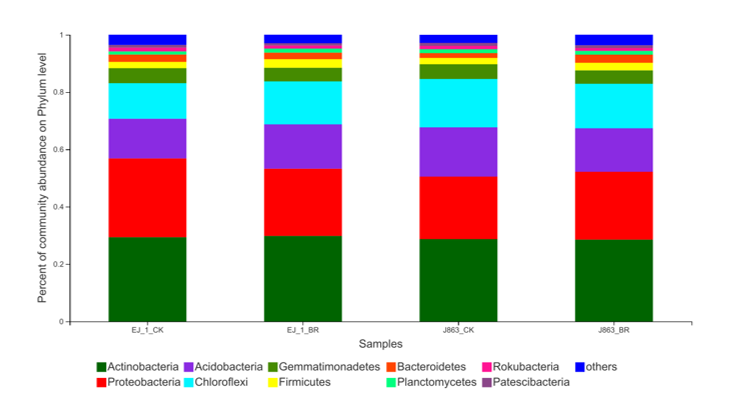
Among all sequences, unknown sequences were classified as “other group”. In Field A, the dominant bacterial phyla were Proteobacteria, Actinobacteria, Acidobacteria, Gemmatimonadetes, Chloroflexi, Bacteroidetes, Planctomycetes, Rokubacteria, Nitrospirae, Verrucomicrobia, Latescibacteria, Firmicutes and Patescibacteria, and these phyla accounted for more than 95% of the total sequences in each sample (Fig. S1). The changes in the relative abundances of the dominant bacterial taxa associated with the different cotton varieties after the application of broccoli residues were compared at the phylum level (Fig. 5). Notably, all dominant bacterial phyla associated with J863 increased in abundance following the application of broccoli residues, while for EJ-1, the dominant bacterial phyla were influenced to different degrees by the application of the broccoli residues. Among them, Gemmatimonadetes, Rokubacteria, Nitrospirae, Verrucomicrobia and Firmicutes increased. The most abundant group was Firmicutes, which increased by approximately 2.36 and 1.41-fold for EJ-1 and J863, respectively, when compared with the values for CK. However, Proteobacteria, Chloroflexi, Bacteroidetes, Planctomycetes, Latescibacteria and Patescibacteria decreased following the application of broccoli residues (Fig. 5A). In Field B, the dominant bacterial phyla were Actinobacteria, Proteobacteria, Acidobacteria, Chloroflexi, Gemmatimonadetes, Firmicutes, Bacteroidetes, Planctomycetes, Rokubacteria, Patescibacteria, Entotheonellaeota, Nitrospirae and Verrucomicrobia, and these phyla accounted for more than 95% of the total sequences in each sample (Fig. S2). For the cultivars EJ-1 and J863, the dominant bacterial phyla were influenced to different degrees by treatment with broccoli residues. Actinobacteria, Gemmatimonadetes and Firmicutes were increased by the application of broccoli residues. The most abundant group was also Firmicutes in Field B, and the fold changes were 1.42 and 1.27 for EJ-1 and J863, respectively. Acidobacteria decreased as a result of the application of broccoli residues. In addition, Proteobacteria, Bacteroidetes, Patescibacteria, Entotheonellaeota, Nitrospirae and Verrucomicrobia decreased for EJ-1, while the opposite tendency was observed for J863 (Fig. 5B). Based on the results for the different cultivars and field sites, the relative abundances of Actinobacteria, Gemmatimonadetes and Firmicutes in the soil increased after the application of broccoli residues.
In Field A, the dominant bacterial genera (cultured and at least one group in two groups average relative abundance > 0.5%) were Sphingomonas, RB41, MND1, Haliangium, Nitrospira, Lysobacter, Bryobacter, Gemmatimonas, Gaiella, Iamia, Pontibacter, Streptomyces, Steroidobacter, Ilumatobacter, Blastococcus, Dongia, Luedemannella, Rubrobacter, Nocardioides, Gemmatirosa, Massilia, Bacillus and Sphingobacterium (Fig. 6A). Among them, the relative abundances of RB41, Gemmatimonas, Pontibacter, Streptomyces, Blastococcus, Massilia and Bacillus were increased by the application of broccoli residues for all cultivars, while decreases in MND1, Nitrospira, Bryobacter, Iamia, Steroidobacter, Ilumatobacter and Dongia were observed after the application of broccoli residues.
In Field B, the dominant bacterial genera were Arthrobacter, Streptomyces, RB41, Rubrobacter, Bacillus, Sphingomonas, Nocardioides, Solirubrobacter, MND1, Gaiella, Blastococcus, Steroidobacter, Bryobacter, Nitrospira, Marmoricola, Microvirga, Microlunatus, Haliangium, Skermanella, Dongia, Intrasporangium, Lysobacter, Gemmatimonas, Novosphingobium, Kribbella and Nitrolancea (Fig. 6B). Among them, the relative abundances of Arthrobacter, Bacillus, Microvirga, Intrasporangium, Lysobacter and Nitrolancea increased, while those of Streptomyces, MND1, Blastococcus, Steroidobacter, Nitrospira, Haliangium, Dongia and Kribbella decreased. Based on the above results at the two field sites, Bacillus was the only genus that steadily increased after the application of broccoli residues.
Relationships between the occurrence of VW and bacterial community composition
The relationships between the occurrence of VW and bacterial community composition in Field A and Field B were studied with RDA (Fig. 7). For Field A, the RDA that was performed with the genera and disease incidence data showed that the first two RDA components could explain 52.3% of the total variation (Fig. 7A). As shown by their close grouping and by the vectors, the disease incidence of cultivar J863 was positively related to the abundant genera Gemmatimonas, Pontibacter, RB41, Blastococcus and Massilia after the application of broccoli residues, and it was negatively related to Bacillus, Lysobacter, and Nitrospira. However, the disease incidence for EJ-1 treated with BR was positively related to the abundant genera Streptomyces, Rubrobacter, Bryobacter and Nocardioides, and it was negatively related to Gemmatimonas, Pontibacter, RB41, Blastococcus and Massilia.
For Field B, the RDA that was performed with the genera and disease incidence data showed that the first two RDA components could explain 47% of the total variation (Fig. 7B). The disease incidence in the BR treatment for the cultivars (including J863 and EJ-1) was positively related to the abundant genera Bacillus, Nocardioides, RB41, Rubrobacter, and Arthrobacter and negatively related to Streptomyces, Nitrospira, Sphingomonas, and Lysobacter.
Control effect of the exogenous application of Bacillus subtilis NCD-2 on VW
The control effect of the application of B. subtilis on cotton VW was investigated in our study. As indicated in Fig. 8, the control effect of BS on the disease at the boll-forming stage for EJ-1 was 38.55%, while that for J863 was 26.73%. Additionally, further study at boll opening showed that the control effect for EJ-1 was 31.96%, while that for J863 was 35.31%. The average control effects for EJ-1 and J863 were 35.26% and 31.02%, respectively.
{kind=link}
{kind=link}