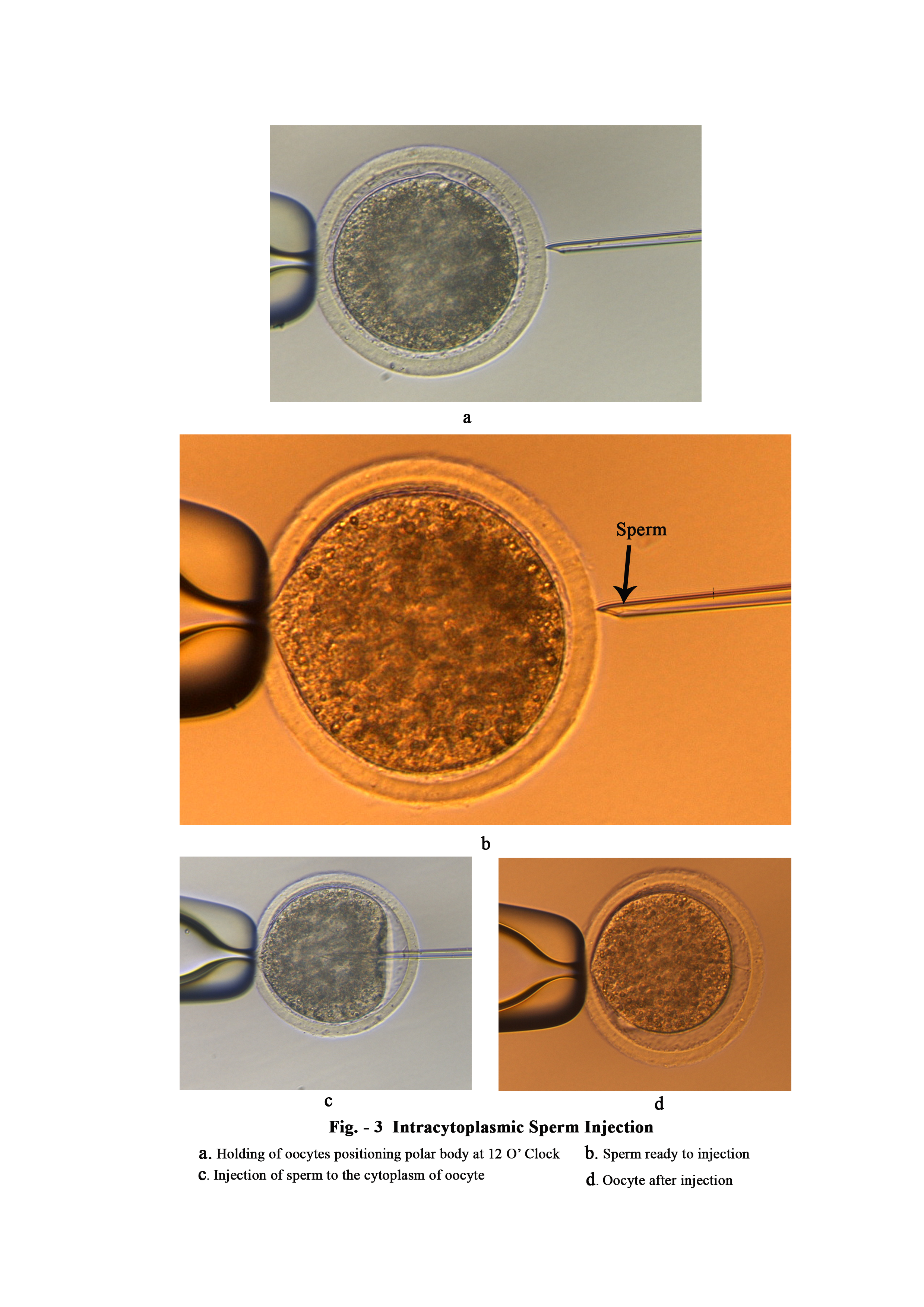
A suitable ICSI protocol for the production of goat embryos from in vitro derived oocytes obtained from slaughterhouse ovaries and epididymal sperm was demonstrated in our study. Sharp injection pipette of 5 µm diameter was found efficient for injection with satisfactory survival rate (83.21%) and lower degeneration rate (17.78%) during in vitro culture. Chemical activation with ionomycin, cyclohexamide and 6-dmap was found satisfactory for oocyte activation and cleavage (63.33%).
One of the important things to initiate the sperm nucleus decondensation and oocyte activation is the mutual interaction of the ooplasmic factors and sperm sub-membrane components (perry et al., 1999a). Permeabilization of the sperm membrane is a prerequisite of this process during normal fertilization (Kasai et al., 1999) or IVF. It is known that a permeabilization treatment of the sperm membrane prior to ICSI helps the process of decondensation of head sperm. In this study, sperm permeabilization was done by tail breaking by injection pipette as describe by Rahman et al. (2007). It was found that it speed up the procedure and more easier than tail cutting. Palermo et al. (1996) found that the fertilization rate of human oocytes following ICSI using testicular spermatozoa increased significantly when sperm tails were scored more harshly by injection pipette. In our experiment the same technique of harsh hitting of spermatozoa tail was applied which resulted in successful embryo production. In this study, immobilized goat sperm cells were used for intracytoplasmic sperm injection to produce good quality embryos. Results indicated that goat sperm motility was not required under the ICSI procedure. This result agrees with results from the human (Palermo et al., 1992) and the cow (Goto et al., 1990). It was also found that the motile spermatozoa after injection keep moving resulted in degeneration of oocytes and did not reached even cleavage stage. The results agree with Kimura and Yanagimachi (1995). Continuous movement of sperm cells within the cytoplasm may disorganize the oocytes structural elements and/or disturb the “oolemma’s wound healing” (Kimura and Yanagimachi, 1995). In the present study it was also observed that 17.78% oocytes degenerate during in vitro culture after sperm injection which may be due to disorganization of oocytes’ structural elements and failure to heal the “oolemmal wound” caused by injection.
Since the oolemma of goat metaphase ii oocytes is characterized by extreme elasticity (wang et al., 2003), like that of bovine oocytes (Gagne et al., 1995), it’s very difficult to broken the oolemma with micropipette insertion. In fact, spermatozoa could have been expelled from the ooplasmic furrow created by the injection pipette. It’s very difficult to inject a sperm into the ooplasm without damaging the oocyte membrane and cytoplasmic structure (Rahman et al., 2007). In our present study 17.78% oocytes damaged due to injection. This may be due to factors mentioned above. To overcome the difficulty of sperm penetration through the oolemma and improve the efficiency of ICSI many workers reported successful application of piezo-drill, a device that generates mechanical pulses through a piezo-actuator to break the oolemma during icsi with a survival rate of 97% (wang et al., 2003). However, sophisticated equipment and extensive manipulating experience are needed to use piezo-driven micromanipulator. Rahman et al. (2007) noticed in his study that the main cause of oocyte degeneration was the failure of the injected oocytes to heal the damage caused by the mechanical insertion of the needle, especially when using a needle of larger diameter without a spike. This problem was minimized by using a needle of 5 µm inner diameter and outer diameter of 7 µm (cook medical, Bloomington, USA). Breaking the oolemma of goat metaphase ii oocyte was confirmed by aspiration of cytoplasm after injection followed by reinjection of the cytoplasmic material. In our present study 83.21% oocyte survived after injection which is lower than piezo-driving actuator (97%) as reported by Wang et al. (2003).
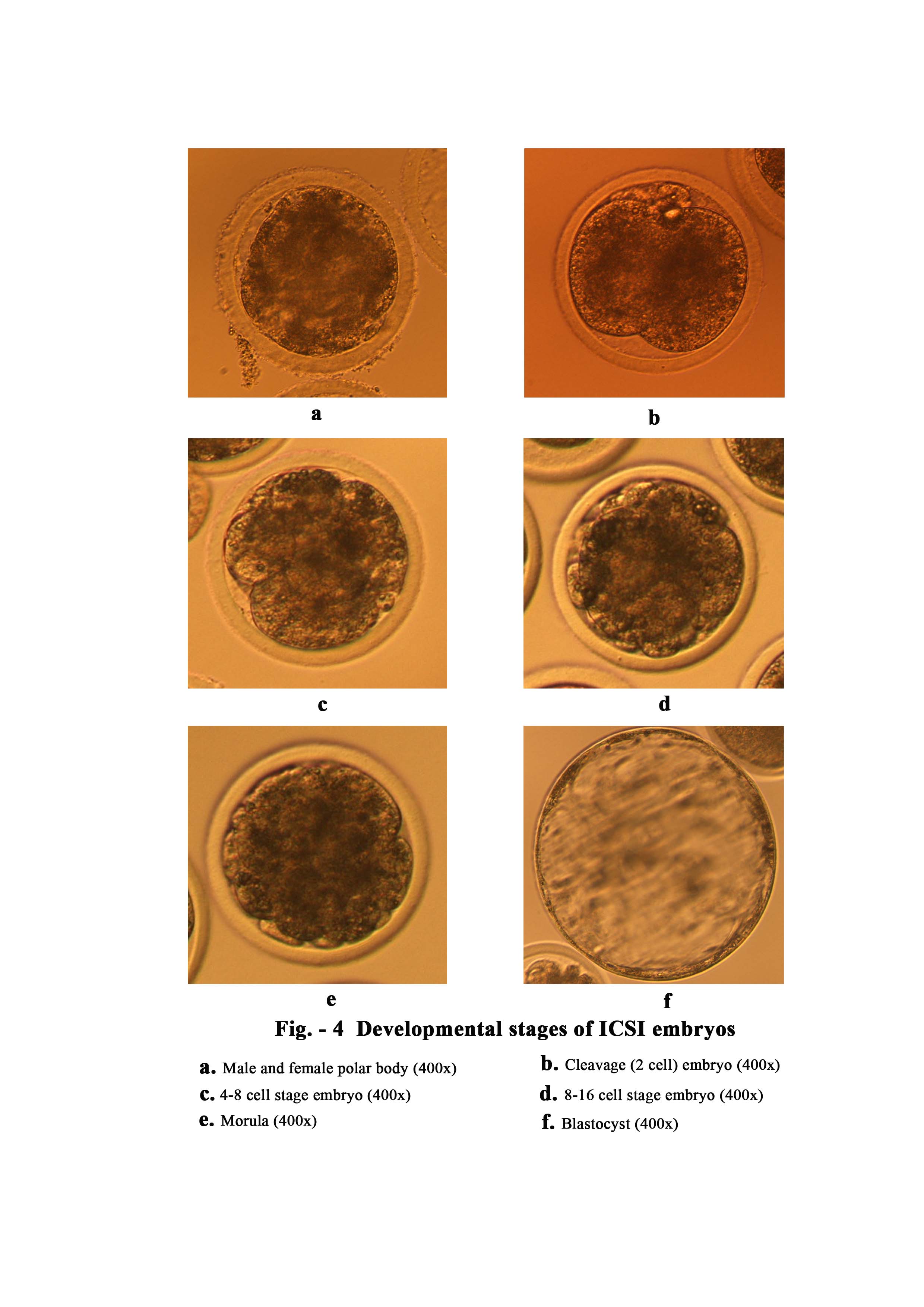
In mice, Kimura and Yangimachi (1995) demonstrated that mouse oocytes injected with testicular and epididymal spermatids or spermatocytes were unable to activate oocytes. Furthermore, oocytes containing injected spermatids or spermatocytes had to be activated artificially to support normal fertilization and full term development. These results demonstrate that artificial stimulation can support full development of mouse oocytes injected with spermatids or spermatocytes. In our study, epididymal sperm where injected for fertilization. Immature spermatids and spermatocytes need chemical activation for further embryonic development. We observed that oocyte activation was more affected by chemical activation than by mechanical sperm injection. Mechanical sperm injection into the ooplasm is enough to activate the oocyte in human (Palermo et al., 1992), hamster (Hoshi et al., 1992) and mouse (Kimura & Yanagimachi, 1995). But, it has been demonstrated that some species, such as cattle (Keefer et al., 1990) and pigs (Proust and Roth, 2003) need the aid of chemical activation after ICSI to improve the embryo development. A number of reports have suggested that acolyte activation could be improved following ICSI with different stimuli like calcium ionosphere in goats (Keskintepe et al., 1997), 6-dimethylaminopurine in cattle (Rho et al., 1998), and electrical pulses in humans (Yanagida et al., 1999) and pigs (Lai et al., 2001). Protocols with the best results in goat were those that combined ionomycin with 6-dmap (Ongeri et al., 2001). In our study, we used conventional manual needle injection, slaughter house derived goat oocytes and epididymal spermatozoa with broken tails, and under these conditions chemical stimulation was needed to achieve the oocyte activation. Ionomycin,CHX and 6-dmap was used to activate injected oocytes. A good percentage (63.33%) of zygotes with male and female polar body was obtained with ICSI plus chemical stimulation. However, 21.11% oocytes were not activated. Jiménez-macedo et al. (2005) reported that 38.6% of ICSI embryos with chemical activation by ionomycin and 6-dmap showed normal fertilization and 42.8% were not activated. In our study activated oocytes were found to be higher (63.33% vs. 38.6%) than reported by Jiménez-macedo et al. (2005) and not activated oocytes were lower (21.11% vs. 42.8%). Artificial oocyte activation always carries the risk of an induction of parthenogenesis. That is why it’s necessary to include sham (injected oocyte with vehicle but without sperm) oocytes as control group of mechanical activation. As expected, out of 22 oocytes no oocytes showed signs of embryonic development.
In order to increase the IVEP efficiency in goat the identification and selection of a suitable system capable of supporting embryonic development in vitro is critical. Using a complex and sequential media consisting of MTALP medium with OGS (20%) for first 24 hours, MKOSM with fatty acid free BSA (1%) from 96–192 hours, Wang et al. (2003) obtained a higher cleavage rate (89%), morula (41%) and blastocyst (21.1%) production. However, using a slightly modified SOF medium, Jimenez- macedo et al. (2005), Jiménez- macedo et al. (2006) and Keskintepe et al. (1997) obtained 53.40%, 66.90% and 62.20% cleavage and 2.7%, 10.60% and 25% blastocyst, respectively. Shadanloo et al. (2010) found 84.80% cleavage and 29.50% blastocyst using immobilized frozen semen after activation with ionomycin. RVCL is a commercial medium from cook’s (Australia), which is basically used for cleavage. However, it has been successfully used for development up to blastocyst stage in buffalo. Shah et al. (2008) used RVCL of cloned embryos and reported significantly higher blast cyst development as compared to mcr2 and msof media. In our study RVCL-blast-BSA media was used as a sequential media. Out of 90 oocytes 67 (74.44%) were cleaved (2 cell), 57 (63.33%) were 4–8 cell, 40 (44.44%) were 8–16 cell, 21 (23.33%) were morula and 13 (14.44%) were blastocyst. Lower percentage of cleavage rate, morula and blastocyst production was obtained than Wang et al. (2003). However, result of the present study is still satisfactory because like Wang et al., (2003) a piezo micromanipulator was not used to facilitate sperm injection. Result of the present study is comparable with Jiménez- macedo et al. (2005) and more than Rahman et al. (2007). Rahman et al. (2007) reported zygote 54.1 ± 5.5%, cleaved and 2–8 cell 18.2 ± 6.9% and 9–16 cell and morula 2.9 ± 2.9%.
Comparison of developmental potency in ICSI and IVF embryo indicated efficiency for in vitro embryo production in goat over ICSI method. Jiménez- macedo et al. (2005) compared developmental potency of goat embryos produced by ICSI and IVF with different culture media and activation protocol. They reported that the percentage of embryo reaching 8–16 cell stage was significantly higher (p < 0.005) in the ICSI than in the IVF group (22.8% and 10.3%, respectively) and the percentage of zygotes was significantly higher (p < 0.05) in ICSI than in IVF group (40% vs. 25.1%). In the present study the percentage of 2 cell stage embryos were significantly higher (p < 0.05) in ICSI group than IVF (73.40 ± 3.12% vs. 64.06 ± 2.44%) in RVCL-blast-BSA media. However, percentage of morula and blastocyst production significantly higher (p < 0.05) in IVF group than ICSI (37.90 ± 3.15% vs. 23.20 ± 2.73% and 21.05 ± 2.09% vs. 14.56 ± 1.75%). The higher percentage of cleavage observed in ICSI oocytes compared to IVF oocytes could be due to: (1) the activation protocol used after ICSI procedure and (2) ICSI allowing the fertilization of all competent oocytes. No significant difference was found in 4–8 cell and 8–16 cell stage embryos.
{kind=link}
{kind=link}